
INTRODUCTION
Bacterial resistance to antibiotics has become one of the major threats to human health worldwide. According to the report released by the UN Interagency Coordination Group (IACG) on Antimicrobial Resistance on 29 April 2019 [1], at least 700,000 people die every year from illnesses caused by drug-resistant bacteria. Immediate action is needed in order to combat the spread of bacterial resistance. Along with the development of new approaches to fighting pathogenic bacteria, the discovery of new potent antimicrobials remains a key path toward this goal.
Antimicrobial peptides represent a diverse class of natural antimicrobial compounds that are produced by invertebrates, plants, and animals in order to protect the host organism from bacteria [2, 3]. These compounds are considered to have potential as new antimicrobial drugs [4, 5]. A number of natural polypeptides that are produced by plants has recently become the subject of intensive research [6-8] marking the beginning of an investigation into this large group of prospective antimicrobial compounds.
According to the structure of these polypeptides, these potential antimicrobial compounds can be divided into cationic, anionic, linear, and cyclic polypeptide groups. Cyclic polypeptides and lipopeptides (cyclic polypeptides with a lipophilic tail) form a special group of compounds that have a number of diverse functions that protect their host organisms in various ways [9]. For example, it is believed that the lipopeptides produced by Pseudomonas and Bacillus act as antiprotozoal, antifungal, antibacterial, and antiviral agents [10]. In the present paper, we will only consider the antimicrobial properties of cyclic polypeptides.
There have been reports that suggest several modes of action of polypeptides as antimicrobial agents: disruption of the bacterial cellular membrane and inhibiting the important live cycle processes in bacteria like the cell wall synthesis, nucleic acid synthesis, and synthesis of proteins [11-13]. The most studied mechanism of action of these antimicrobial compounds is the disruption of the bacterial cell membrane that leads to the efflux of metal cations and/or molecules from the bacterial cell and eventually cause the death of bacteria [12, 14]. This unique mode of action gives these compounds a number of advantages compared to the antibiotics of the other classes:
They are active against a wide variety of Gram-positive and Gram-negative bacteria.
They do not need to be transported inside the cell of Gram-positive bacteria because they interact with the bacterial membrane from the outside. In case of Gram-negative bacteria, the interaction with antimicrobial peptides is more complex since these bacteria have both inner and outer membranes.
Some of the most abundant mechanisms of bacterial resistance (e.g. multidrug efflux pumps) do not affect polypeptides since they act from outside of the bacterial membrane.
Over the course of studies of the antimicrobial action of linear polypeptides, four general mechanisms of bacterial membrane disruption by these compounds have been suggested: the formation of ‘barrel-stave’-like pore, of a toroidal pore, the ‘carpet’ mechanism, and a ‘detergent-like’ mechanism [12, 14, 17, 18]. The mechanism of interaction of antimicrobial polypeptides with the bacterial membrane could change depending on a number of conditions, e.g. on the local concentration of the corresponding polypeptide at the bacterial membrane surface.
HYPOTHESIS
Cyclic polypeptides and lipopeptides
Cyclic polypeptides (as well as lipopeptides) have particular physico-chemical properties that determine the unique mode of their interaction with the bacterial cell membrane.
There are several examples of lipopeptides – daptomycin [19] (Fig. 1), colistin (polymyxin E) [20], and polymyxin B [21] (Fig. 2) – and cyclic polypeptides – tyrocidine and gramicidin S (Fig. 3) [22] – that are used or were used as antibiotics in clinical practice.


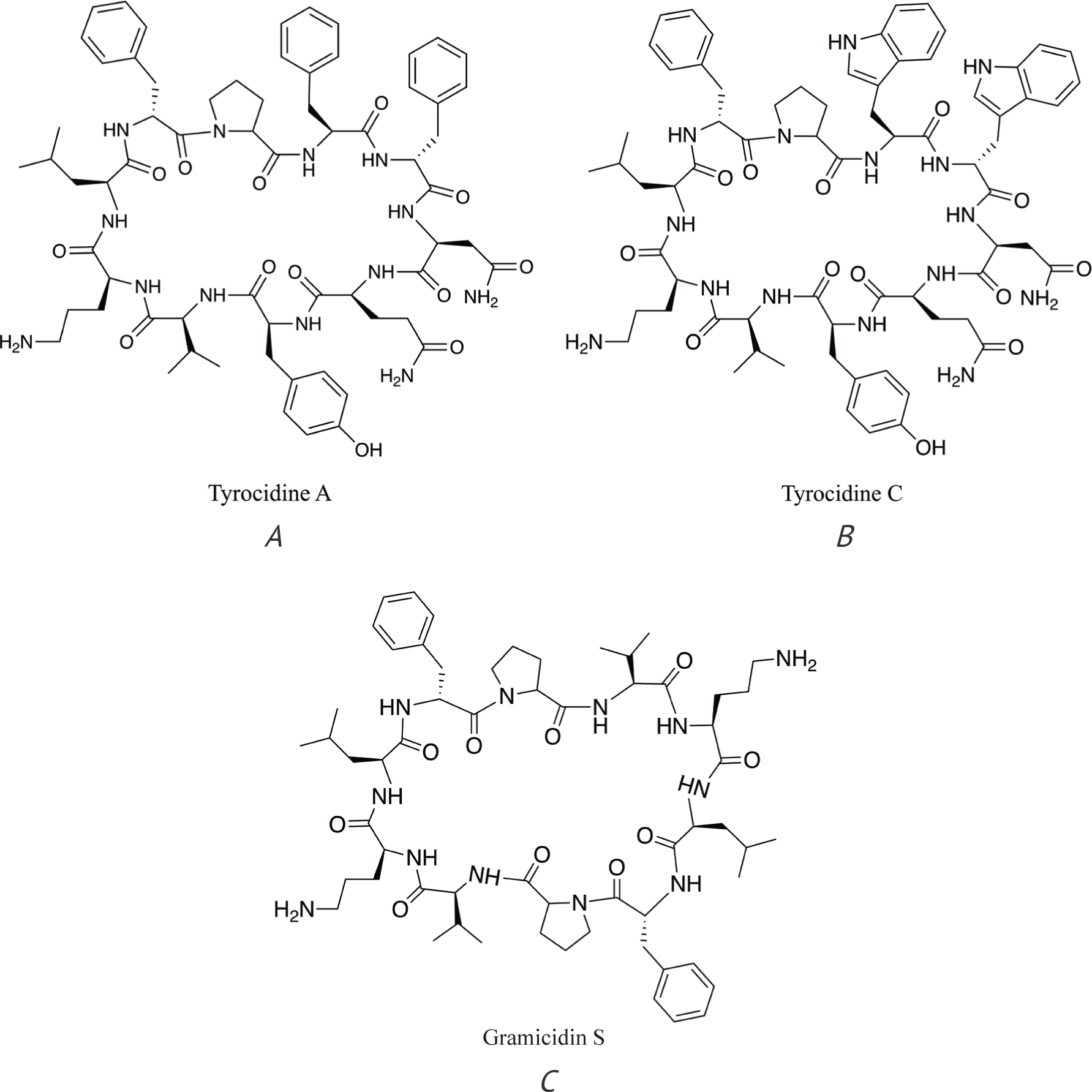
Tyrocidine and gramicidin S have strong hemolytic potential and, therefore, their use is limited to topical application. Both colistin (polymyxin E) and polymyxin B (Poly-Rx) are active against Gram-negative bacteria. The use of colistin was practically discontinued in the 1980s due to the side effects but since the 1990s, it is back in use in clinical practice. Daptomycin is effective against Gram-positive pathogens, including a number of otherwise resistant bacterial strains, such as methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-intermediate Staphylococcus aureus (VISA), and vancomycin-resistant Staphylococcus aureus (VRSA) as well as several Streptococcus and Enterococcus species, including vancomycin-resistant Enterococcus bacteria (VRE) [19, 23]. Daptomycin is used in clinical practice for the treatment of complicated skin infections, bacteremia, and right-sided endocarditis caused by Gram-positive bacteria.
Daptomycin mechanism of action
Since daptomycin is the only lipopeptide antibiotic that is widely used in clinical practice and considered as the drug of choice against methicillin-resistant Gram-positive pathogens (along with vancomycin and linezolid), there have been a number of studies focused on the mechanism of action of this antibiotic [23-28].
Daptomycin is a cyclic lipopeptide produced by Streptomyces roseosporus using non-ribosomal peptide synthetases. It was discovered by Eli Lilly and then studied and commercialized by Cubist Pharmaceuticals in 2003 [28]. Daptomycin consists of 13 amino acids: 10 amino acids (L-Gly, L-Orn, L-Asp, D-Ala, L-Asp, L-Gly, D-Ser, L-threo-MeGlu, L-Kyn, L-Thr) form a ring that is closed by an ester bond and the other 3 amino-acids (L-Asp, D-Asn and L-Trp) are in the side chain with a terminal decanoic acid residue (Fig. 1) [28, 29]. Since daptomycin contains four carboxylic groups that are dissociated at the physiological pH and one primary amino group that is protonated at physiological pH, the total charge of this molecule is -3 (Fig. 1). It was shown in a number of studies that the mode of daptomycin action involves the disruption of the bacterial cell membrane [24, 26, 28-30]. However, other possible mechanisms of action of this antibiotic have been discussed in the literature, including lipid extraction [31], disruption of bacterial cell wall synthesis, and interaction with membrane-bound proteins [23, 32, 33]. Since the permeabilization and depolarization of the bacterial cell membranes containing a high content of negatively charged molecules of phosphatidylglycerol (PG) and cardiolipin (CD) by daptomycin is well documented [24, 26, 28-30], it should be considered as the main mechanism of action (or possibly a necessary step in the multistep mechanism of action) of this antibiotic against the corresponding bacteria. The fact that bacterial strains containing much less or no anionic phosphatidylglycerol in their membranes are resistant to daptomycin [25, 28, 34, 35] serves as additional evidence that the main mechanism of action of daptomycin is bacterial membrane disruption. However, it does not mean that this is the only mode of action of this antibiotic against bacteria. Since a number of different methods (recently review by Raheem and Straus [17]) are used to study the interaction of antimicrobial peptides (AMP) with biological and model membranes (including the formation and function of ion channels), there are very different opinions in the literature about the mechanism of action of the same AMPs including ionophores.
Thus, there is an opinion that daptomycin does not form ion channels in bacterial (or model) membranes [32, 33, 36]. However, this opinion was challenged in the recent excellent paper by Seydlova et al. [37]. Along with important experimental evidence of the pore-forming ability of daptomycin in model and bacterial membranes, the authors of this publication discuss the possible reasons for several negative results of analogous experiments published elsewhere. One of the main arguments supporting the statement that daptomycin does not form pores (or ion channels) in bacterial membranes is the observation that this antibiotic is significantly less effective in causing the leakage of K+ or other cations from the bacterial cell or model membranes than known ionophores like valinomycin and gramicidin [32, 35, 36]. That argument should be considered with caution because membrane permeabilization by daptomycin significantly depends on the membrane potential [37] and also because ionophores like valinomycin and gramicidin have different mechanisms of action (Fig. 4, Fig. 6B).

Valinomycin is known to efficiently transport potassium ions across membranes, including bacterial cell membranes, and is considered as a highly toxic compound. Valinomycin is a 12-unit depsipeptide that contains alternating D- and L-valine, D-hydroxyisovaleric acid, and L- lactic acid residues that are connected by amide and ester bonds (Fig. 5). Since valinomycin has a hydrophilic cavity made by 12 carbonyl groups, it readily forms a complex with potassium ion. This complex is efficiently transported through the lipophilic membrane because the iso-propyl residues pointing outside the cavity smoothly interact with the membrane components. The ionophore molecule travels back and forth through the membrane repeatedly, each time transporting a single K+ ion from the cytoplasm of bacterial cell to the extracellular space [38-40] (Fig. 4).

Gramicidin is a mixture of 3 linear 15 amino acid non-ribosomal polypeptides that have different amino acids in position 11 and can have a different amino acid at position 1 (Fig. 6A). In membranes, it can adopt the beta-helical conformation or double helical structures [41].

When two beta-helical conformations in both membrane leaflets form a dimer, the transmembrane ion channel is open and monovalent ions (e.g. Na+ or K+) can freely travel through the ion channel [41] (Fig. 6B).
Gramicidin (Fig. 6) forms very efficient ion channels. According to Gumila et al. [42], gramicidin channels are much more effective in ion transport through the phospholipid membrane of erythrocytes than the well-known ionophores valinomycin (~150 times) and nonactin (Fig. 7) (~12,000 times).

It is also approx. 19,000 times more effective than gramicidin S, which is a cyclic polypeptide and contains 10 amino acids in its cycle (Fig. 3), although there is no direct evidence that gramicidin S forms ion pores in the bacterial membrane. However, its structure is very similar to that of tyrocidine A (Fig. 3) that is known to form ion channels in membranes, and both gramicidin S and tyrocidine A have similar activity against pathogenic bacteria [22]. The interaction of gramicidin S with the bacterial membrane leads to membrane depolarization, and it has recently been proposed to form ‘transient pore-like zones’ in membranes [43]. It should be mentioned that both gramicidin S and tyrocidine A have other modes of action, e.g. delocalization of membrane proteins involved in cell division [22], but it could be one of the consequences of membrane depolarization. It should also be mentioned that 8-10 molecules of daptomycin (as will be discussed further) as well as the presence of Ca2+ are necessary for the formation of oligomer that causes potassium ions efflux. This could be one of the reasons for the significantly lower activity of daptomycin in experiments involving ion transport through the membrane compared to active ionophores.
The determination of the exact structure of daptomycin oligomer is extremely important because it will lead to a much better understanding of the interaction of daptomycin molecules with the bacterial cell membranes. This, in turn, will help to design new effective antibiotics based on the daptomycin structure.
The efforts of several research groups were concentrated on a detailed study of the mechanism of bacterial cell membrane disruption by daptomycin [24, 26-30, 44]. These studies proved that the interaction of daptomycin with the bacterial cell membrane causes potassium efflux and membrane depolarization, which eventually leads to bacterial death, probably as a result of membrane disruption or more complex processes involving the membrane restructuring and/or dislocating of membrane-bound proteins. Daptomycin does not form large pores in the bacterial membrane [37, 45, 46]. Pores that are formed by daptomycin oligomers in the bacterial membrane are specific for the efflux of sodium and potassium cations [45, 46], but other small cations (e.g. Mg2+) are also transported. In studying the daptomycin mechanism of action, it was shown that the presence of Ca2+ is necessary for the interaction of daptomycin with bacterial cell membranes as well as with model membranes containing negatively charged phosphatidylglycerol (PG) and/or cardiolipin (CD) [24, 27, 47, 48]. As a result of these studies, scientists suggested two slightly different mechanisms of membrane permeabilization by daptomycin that include a three step scheme (Fig. 8) [24] or a four step scheme, where steps 1-2 are binding and the insertion of daptomycin in the membrane and steps 3-4 are oligomerization and pore formation [27]. On the other hand, Ming-Tao Lee et al. [49] suggested that there are only two steps: step 1 – interaction of daptomycin with the bacterial membrane – and step 2 – its oligomerization in the membrane. However, all of the scientists who have studied the mechanism of action of this antibiotic agree that the oligomerization of daptomycin in the bacterial membrane is essential for killing bacteria [24, 27, 29]. According to the most accepted mechanism of bacterial membrane disruption, the first step is the initial interaction of daptomycin with the bacterial cell membrane (Fig. 8). Since both daptomycin and the bacterial membrane, which contain phosphatidylglycerol (PG), are negatively charged, it is believed that the initial interaction of the lipopeptide molecule with the membrane is accomplished through coordination with calcium ions [24].

Recent studies showed that daptomycin forms a complex with Ca2+ cation and phosphatidylglycerol on the surface of membrane before insertion into the membrane [35]. That can serve as evidence of the important role of Ca2+ in the interaction of daptomycin with the bacterial membrane. The next step is the insertion of daptomycin (deeper) into the membrane followed in the last step by the oligomerization of lipopeptide in the bacterial membrane [26, 29, 44, 50]. It is noteworthy that oligomerization in membranes was also observed for daptomycin analogue lipopeptide antibiotic A54145 [51]. It is believed that the oligomers of daptomycin make pores in the bacterial membrane that thereby causes the efflux of potassium ions from the bacterial cell and membrane depolarization leading to bacterial death [24, 28, 37]. The exact structure of the daptomycin oligomers in the membrane is still unknown and remains an objective of experimental studies as well as a subject of ongoing discussion in the scientific literature. Several studies have been focused on the detailed evaluation of daptomycin oligomerization in the bacterial membrane. In one study by Muraih and Palmer [30], the number of daptomycin molecules in the oligomer was determined using fluorescence resonance energy transfer (FRET). According to their results, the membrane-bound oligomer contains 6-7 molecules or possibly twice as many if the oligomer extends through both membrane leaflets. More recent studies showed that four molecules of lipopeptide are located in the cytoplasmic membrane leaflet while the other four molecules are in the exoplasmic leaflet (Fig. 9) and, therefore, a total 8 molecules of daptomycin are needed for the formation of the pore (or ion channel) in the membrane [28, 52, 53]. Seydlova et al. [37] observed variations in conductivity consistent with a variable number of channel subunits with the most abundant channels formed by 4 or 5 subunits. These scientists then sought to reconcile their findings with those of Zhang et al. [52], who conducted experiments in the absence of a membrane potential, by postulating that a second oligomer of the same size could be present in the opposite membrane leaflet. While this may be so, we should also consider the possibility that, in the presence of a high membrane potential, a single tetramer or pentamer may be sufficient to permeabilize the membrane.

It should be mentioned that the same number of oligomer subunits was proposed for the daptomycin analogue lipopeptide antibiotic A54145 [29].
Self-assembling cyclic peptides
On the other hand, self-assembling cyclic peptide nanotubes (or nanowires) that can function as ion channels in membranes has been well documented. In the last 10 years, there has been a number of research papers and reviews that describe and discuss this phenomenon, e.g. [54-59]. According to these data, over the course of the interaction with biological and model membranes, cyclic peptides spontaneously form nanotubes or nanowires – structured aggregates or oligomers. These nanotubes are formed by self-assembling of cyclic peptides and can serve as efficient ion channels in phospholipid membranes. A number of publications, for example [60-63], describe the spontaneous formation of dimers and nanotubes (or nanowires) by cyclic polypeptides in organic and aqueous solutions with a different pH. A review by Rodriguez-Vazquez [64] describes the recent advances in studies of the self-assembling of cyclic peptides in membranes. This paper covers a number of experimental results that showed that the interaction of cyclic peptides with membranes leads to the spontaneous formation of nanotubes that function as ion channels. The cyclic part of the molecule in these polypeptides is usually formed by 8-12 alternating D- and L-amino acids, although cyclic peptides with a different structure and cyclic urea derivatives also form nanotubes [64, 65]. Some of these molecules have ‘tails’ containing hydrophilic and/or lipophilic fragments. Tryptophan residues are considered ideal for the integration of the nanotube into the bilayer membrane [64, 66] (Fig. 10).

It was proved that the nanotube formed by self-assembling of 8 molecules of cyclic polypeptide is long enough to span a lipid bilayer and function as a transmembrane channel [57, 66, 67] (Fig. 11).
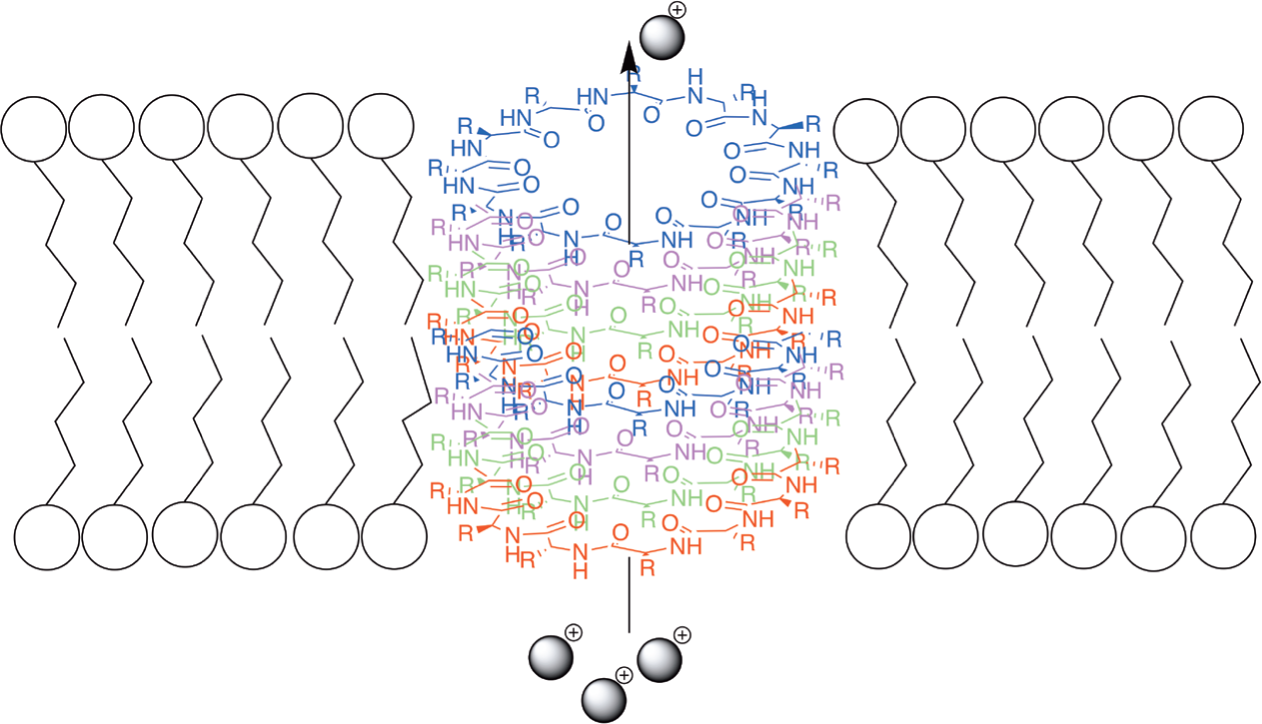
According to Rho et al., cyclic peptides with longer side chains form even more stable aggregates (nanowires) in aqueous solutions [59]. Based on the extensive study of self-assembling cyclic peptides, Rodriguez-Vazquez et al. concluded that cyclic polypeptides with hydrophobic side chains form nanotubes oriented perpendicular to the membrane [64]. On the other hand, Motiei et al. [68] described the synthesis and antibacterial activity of novel self-assembling cyclic D,L,-α-glycopeptides (e.g. Fig. 12) and proved the membrane permeabilization mode of action of these compounds.

Since the formation of these nanotubes in phospholipid membranes is a spontaneous process and the change in Gibbs free energy in the system is negative, this process is exergonic [69]. This means that nanotubes are that particular structure of the cyclic polypeptide oligomers that brings the interaction energy of these oligomers with the membrane to a minimum and, therefore, is the most stable conformation for the membrane – polypeptide oligomers system. Accordingly, and also taking into account the large amount of experimental data on cyclic peptide nanotubes and the close structure of the cyclic daptomycin fragment to the structure of efficient ionophores like valinomycin (Fig. 5) as well as the experimentally determined number of the daptomycin molecules in oligomer – 8, we come to conclusion that the probable structure of daptomycin oligomer in the bacterial membrane is the nanotube that functions as an ion channel (Fig. 13).
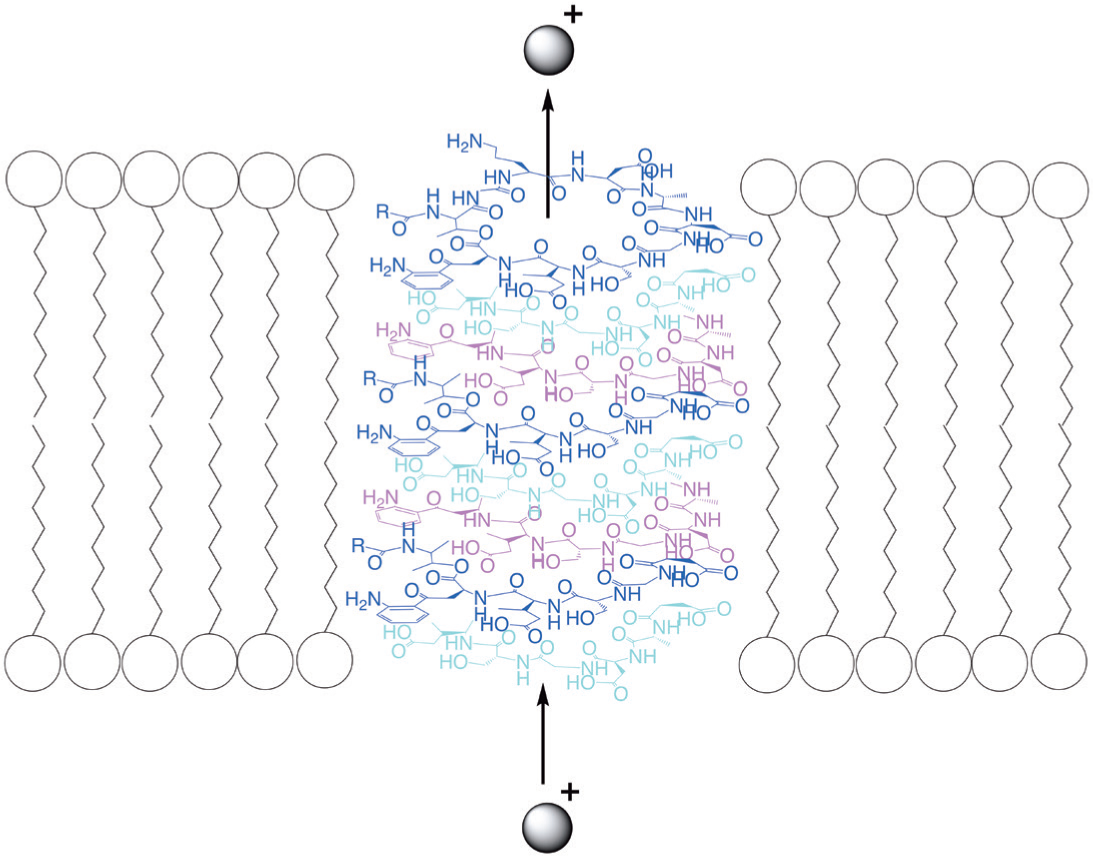
It should be mentioned that, based on an excimer fluorescence study of perylene substituted daptomycin, Muraih et al. suggested that ‘neighboring oligomer subunits are aligned in parallel fashion or form an acute angle with one another’ [50]. Although the self-assembling of cyclic polypeptides into nanotubes is spontaneous, the polypeptide molecules in solution are always in dynamic equilibrium with the nanotube structures in the membrane. This leads to the formation of nanotubes containing fewer molecules than is required to span a lipid bilayer. These nanotubes will function as much less effective ion channels [64, 66].
Considering the interaction between daptomycin molecules over the course of stacking into the nanotube could also help explain the change of the antibiotic activity as a result of the certain modification of its structure.
Attempts to modify the structure of daptomycin showed that changing one (or even two) particular amino acids in the cyclic fragment of the molecule does not cause significant changes in activity [70] and that, occasionally, small changes – like the methylation of amino group in kynurenine – can lead to the formation of more active compounds [71, 72]. On the other hand, activity was lost as a result of the modification of different amino acids [72, 73]. Moreira et al. showed that the chirality of the molecule is extremely important for antimicrobial activity [74].
It is possible that these results could be better explained by taking into account the interaction of daptomycin molecules with each other over the course of stacking into the nanotube where, depending on the specific interaction between the molecules, some functional groups are much more important than others. Chirality is also very important since it determines the interaction of cyclic polypeptides over the course of self-assembling into a nanotube [57, 58, 64]. On the other hand, the introduction of different bulky lipophilic substituents in ornithine residue by reductive alkylation did not significantly change the activity of daptomycin [75]. Since the ornithine amino group is pointing away from the cyclic fragment of the molecule, substituents at this position will not intervene in the stacking process. On the contrary, these substituents will interact with the lipophilic part of membrane, which should increase the stability of the nanotube in the membrane. In fact, daptomycin, with highly lipophilic 4-(phenyl)benzyl fragment in ornithine fragment, showed enhanced activity against S. aureas in the absence of serum [75].
It is possible that the interaction of daptomycin molecules with Ca2+ ions is one of the driving forces for the self-assembling process [29] as it is described for other divalent cations for the self-assembling of helical peptides into nanostructures [55].
FUTURE WORK
The hypothesis that daptomycin forms self-assembled nanotubes in membranes needs experimental validation. We suggest here the possible directions for the experimental and modeling studies that could help prove or disprove this hypothesis:
Determine if daptomycin oligomer (or nanotube) in the membrane contains calcium ions.
Conduct a molecular modeling study on the interaction of daptomycin molecules within the nanotube based on the results of the previous experiment. Determine which daptomycin functional groups are critical for the formation of the nanotube.
Synthesize daptomycin analogs with one or several changed critical functional groups and study the oligomerization of these analogs in membranes and/or their antimicrobial activity.
CONCLUSION
If we consider the formation of nanotubes that function as ion channels in the bacterial membrane as the major mechanism of action of the cyclic polypeptides and lipopeptides, we will have to consider the interaction of these molecules with each other and with the cell membrane over the course of the nanotube formation. This will lead to a better understanding of the structure – activity relationship of cyclic polypeptide and lipopeptide antibiotics – and will help better explain the published experimental data on the in vivo activity of the different compounds of this class. On the other hand, a better understanding of the nanotube structure, formed by daptomycin, that functions as an ion channel in bacterial membranes, will also lead to the successful design and synthesis of new active lipopeptide antibiotics.