
INTRODUCTION
Influenza viruses with shortened nonstructural NS1 proteins have reduced reproductive activity in the respiratory tract of the host organism but can cause comprehensive humoral and cellular immune responses. That enables the development of live attenuated influenza vaccines based on this kind of viruses [1, 2]. The formation of virus-specific CD4+ and CD8+ T-lymphocytes in lungs and in lung draining lymph nodes as well as the significant antibody responses to the surface antigens of the influenza viruses with the modified NS genes has been shown in a number of studies: in mice [3, 4], horses [5], pigs [6, 7], poultry [1], ferrets [8], and macaques [9]. A live intranasal influenza vaccine, which was developed on the basis of the influenza virus with the removed NS1 protein reading frame, has successfully passed clinical trials [2]. The modification of the NS gene by the introduction of the foreign genome fragments from different pathogens is a promising approach for the creation of mucosal vector vaccines against various infections. In particular, immunization with a strain that contains NS1 gene expressing Mycobacterium tuberculosis ESAT-6 protein results in the formation of a protective ESAT-6-specific Th1-type immune response [10].
Despite numerous studies conducted on the impact of the NS1 protein modifications on the formation of adaptive immunity to the influenza virus, the role of the cell-mediated innate immunity on the early stages of the immune response is still poorly understood. The cells of innate immunity serve as the first line of the body’s immune defense against viral infections. These cells are responsible for the activation of adaptive immunity through the processing and presentation of antigens in the context of the major histocompatibility complex (MHC) as well as for the production of proinflammatory and regulatory cytokines, chemokines, and differentiation factors [11, 12].
It is known that the full-length NS1 protein is directly involved in inhibiting the reactions of the innate immune response against the influenza virus by suppressing the expression of genes of type I interferons (IFNs) and other proinflammatory factors [13]. In addition, the action of the NS1 protein has an impact on the effectiveness of the presentation of viral antigens by MHC molecules and, as a consequence, on the immunogenicity of influenza virus proteins [13, 14].
The goal of this study was to compare the cytokine production, kinetics of innate immune cell populations, expression of activation markers in antigen presenting cells (APC), and the levels of influenza-specific T-cellular response in mice immunized with virus containing NS1 protein shortened to 124 amino acid (a.a.) residues (A/PR8/NS124 or NS124) versus mice immunized with the virus with full length NS1 protein (A/PR8/full NS or full NS).
Taking into account the different reproductive activity of viruses with the truncated and intact NS1 proteins in the respiratory tract of animals, an intraperitoneal route of immunization was chosen in this study. This approach allowed for the compensation of the potential differences in antigenic load since the influenza virus is unable to reproduce in the peritoneal cavity [15]. The intraperitoneal immunization of mice with viruses containing full-length and modified NS1 proteins was described by B. Ferko et al. [4] when they used this approach to evaluate the production of cytokines by different viruses.
MATERIALS AND METHODS
Viruses
Two strains based on the influenza A/Puerto Rico/8/1934 (H1N1) virus (A/PR8) encoding (1) the full-length NS1 protein (A/PR8/full NS) and (2) shortened to 124 a.a. NS1 protein (A/PR8/NS124) described earlier [3] were used in this study. The viruses were produced in developing chick embryos (CEs) and purified by high-speed centrifugation in a sucrose density gradient by the standard procedure.
Laboratory animals
The study was performed on C57BL/6 mice obtained from the Biomedical Science Center (Stolbovaya, Russia). All of the experiments in this research study were conducted according to the guidelines for care and work with laboratory animals [16].
Determination of the virus infectious activity
The infectious activity of the virus strains was estimated by a limiting dilution assay performed in a chicken embryo kidney (CEK) cells cultivated in DMEM-F12 medium (Gibco, USA) containing 6 mM GlutaMAX (Gibco, USA), 1 mM sodium pyruvate CH3(CO)COONa (Gibco, USA), and 1% antibiotic-antimycotic solution for cell cultures
(amphotericin B, penicillin, streptomycin; Gibco, USA). The assay was performed in 96-well culture plates (Nunc, Denmark) by the addition of 100 μl of prepared dilutions of the virus-containing material into the wells, and subsequent incubation for 5 days at 34°C and 5% CO2. The results were evaluated visually by cytopathic effect estimation, and a hemagglutination reaction (using a 0.5% suspension of chicken red blood cells) was used as the control. The calculation of virus 50% tissue culture infectious dose (TCID50) was performed using the method described by Reed and Munch [17], and the viral titer was expressed as log10TCID50/ml.
In order to determine the dynamic of weight change, and the reproductive activity of the virus strains in the respiratory tract of mice, animals were immunized intranasally (i.n.) with 30 μl of a viral suspension (6.0 log10TCID50/mouse) under light ether anesthesia. Lung collection was performed on the 2nd, 4th, and 6th days after infection. Toward this end, the mice were sacrificed by cervical dislocation, after which the lungs were removed. The organs were homogenized using a Tissue Lyser II device (Qiagen, Germany) at 30 Hz for 6 min. The viral loads in the lungs of the immunized animals were estimated by the titration of tissue homogenates in the Madin-Darby Canine Kidney (MDCK) cells and expressed as log10TCID50/ml.
Immunization of the animals
The mice were immunized intraperitoneally with a suspension of the corresponding virus in 500 μl of phosphate buffered saline (PBS, Biolot, Russia) solution (7.1 log10TCID50/mouse). The animals of the control group received the same volume of pure PBS.
Analysis of cytokine production in peritoneal washings
In order to collect peritoneal washings, the mice were sacrificed by cervical dislocation 12 h after immunization and 1 ml of cold PBS was injected into the abdominal cavity using a syringe. After 5 min, the peritoneal washings were collected and transferred to test tubes. Samples were centrifuged for 10 min at 500 g (4°C) in an Eppendorf 5810R centrifuge (A-4-81 rotor). The concentration of inflammatory cytokines in the supernatants was determined using the LEGENDplex Mouse Inflammation Panel Kit in V-bottom plates (Biolegend, USA) according to the manufacturer’s instructions.
Flow cytometry
In order to analyze the dynamics of innate immunity cell populations, peritoneal washings were obtained 12 h and 24 h after immunization. Cells (106 in 100 μl) were stained with fluorochrome-conjugated antibodies. Spleen extraction and splenocyte isolation were performed at 8 and 21 days post immunization (d.p.i.) to analyze the adaptive immune response. Cell stimulation was performed in flat-bottomed 96-well plates (Nunc, Denmark) for 6 h with a mixture of the ASNENMETM peptide and brefeldin-A (BD Biosciences, USA). The ASNENMETM peptide corresponds to the highly immunogenic epitope of the influenza A/PR8 NP protein. Two panels of fluorochrome-conjugated antibodies (BioLegend, USA) were used to identify the innate immunity cell populations and to analyze the activation marker expression levels: 1) CD11b-PE/Cy7, CD11c-PE, MHCII-Alexa488, CD103-PerCP-Cy5.5, CD45-APC/Cy7, CD64-BV421, CD24-BV510; and 2) CD45-APC/Cy7, MHCII-Alexa488, Ly6G-PerCP-Cy5.5, CD86-BV421, and CD83-BV510. Identification of the adaptive immunity cells was performed using the fluorochrome-conjugated antibody set containing CD4-PerCP-Cy5.5, CD8-PE/Cy7, CD62L-APC/Cy7, and CD44-BV421 (BioLegend, USA). Intracellular production of cytokines was assessed using antibodies against IFNγ-FITC, IL2-PE, and TNFα-BV510 (BioLegend, USA).
Staining for the detection of intracellular markers was performed using the Fixation and Permeabilization Solution reagent kit (BD Biosciences, USA) according to the manufacturer’s instructions. Zombie Red viability marker (BioLegend, USA) was used to identify the dead cells. True Stain reagent, containing antibodies to CD16/CD32, was used to block non-specific antibody binding (BioLegend, USA). Data were collected on a BD FACSCanto II flow cytometer (BD Biosciences, USA). The results were analyzed using the Kaluza Analysis 1.5a program (Beckman Coulter, USA).
RESULTS
Pathogenicity of virus strains in mice and their reproductive activity in the respiratory tract of animals
The effect of shortening of the NS1 protein on the pathogenicity and reproductive activity of the influenza virus was evaluated by a comparison of the changes in the body weight of mice immunized with A/PR8/NS124 and A/PR8/full NS viruses. The single immunization of the animals with the virus A/PR8/NS124 containing the shortened NS1 protein did not lead to a decrease in body weight (Fig. 1). However, the immunization of animals with an equivalent dose of the pathogenic A/PR8/full NS virus led to a drop in the total body weight of the group to 85% of original weight by the 6th day after infection.
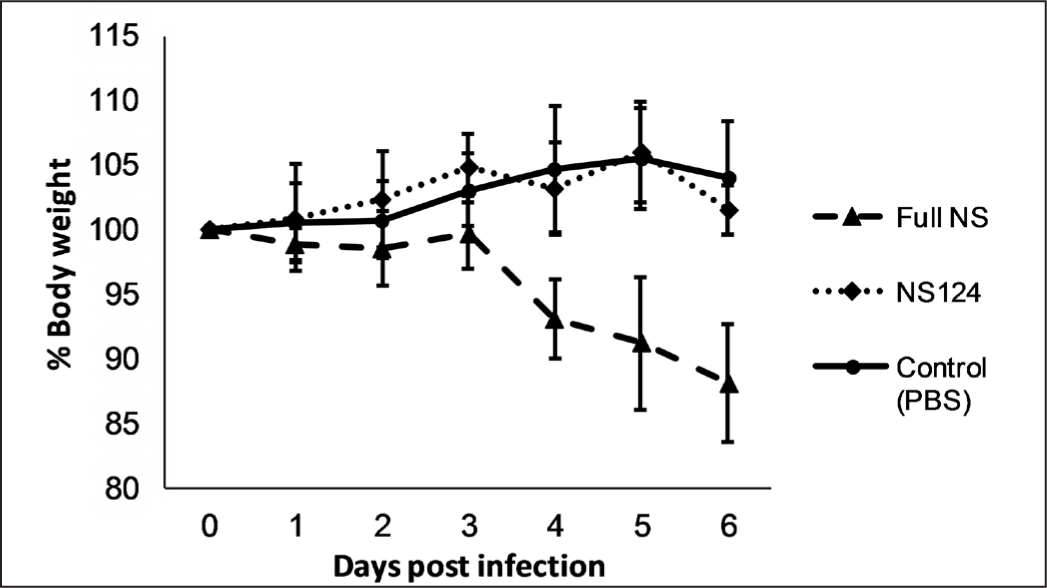
The data are presented as a mean of the percentage of the body weight change from the initial weight value (Mean±SEM). Each experimental group included 10 mice (n=10). The number of mice in the control group: n=6.
In order to determine the reproductive activity of the virus strains, viral loads in the lungs of immunized animals were estimated by the titration of tissue homogenates in the MDCK cells. The results are shown in Table 1.
| Virus strain | Viral titer in the lungs of mice (log10TCID50/ml) | ||
|---|---|---|---|
| 2nd day | 4th day | 6th day | |
| A/PR8/NS124 | 3.70 ± 0.71*** | 3.80 ± 1.20*** | 4.40 ± 1.85 |
| A/PR8/full NS | 6.80 ± 0.71 | 7.00 ± 0.54 | 6.50 ± 0.67 |
Two groups of mice were immunized i.n. with A/PR8/full NS and A/PR8/NS124 viruses (6.0 log10TCID50/mouse). On the 2nd, 4th, and 6th days, 5 animals from each group were sacrificed and the viral titers in the lungs were determined. The data are presented as Mean ± SD.
indicates the significant difference (p<0.001, Student’s t-test) between the groups.
The obtained results show that the shortening of the NS1 protein leads to the attenuation of the virus, which is proved by the lower reproduction rate of the virus in lungs and by the absence of the body weight loss of the immunized animals. Statistically significant differences in viral titers were observed on the 2nd and 4th d.p.i. (p=0.0008 and p=0.003, respectively), whereas on the 6th day the difference between the groups was not significant (p=0.08).
Increased cytokine induction by A/PR8/NS124 versus A/PR8/full NS strain
Considering the differences found in reproductive activity between the A/PR8/full NS and A/PR8/NS124 strains in the respiratory tract of mice, an intraperitoneal route of immunization was chosen in order to compare the immunogenicity of these viruses.
The analysis of the concentration of inflammatory cytokines in peritoneal washings of mice showed that immunization with the A/PR8/NS124 strain leads to an increased induction of cytokines compared to the virus with the full-length NS1 protein (Fig. 2). The most pronounced difference was observed for IFNβ: the production of this cytokine by mice in the NS124 group was almost 300 times higher than by animals in the full NS group (65635.7±14650.3 pg/ml and 223.6±140.3 pg/ml, respectively). In addition, a significant difference was detected in the IL27 levels: the concentration of this cytokine in the peritoneal washings of mice immunized with the virus containing truncated NS1 protein was fourfold higher than that in the group immunized with the full-length NS1 protein (508.6±193.5 pg/ml and 125.2±64.6 pg/ml, respectively). The levels of IL6 and MCP1 production increased in both groups following the immunization. At the same time, in the animals immunized with the A/PR8/NS124 virus, these levels were on average 1.8 (IL6) and 2.0 (MCP1) times higher than in mice that received the A/PR8/full NS strain. The same pattern was observed for the TNFα, IL2p70, and IL1β cytokine concentrations which were 1.5-2.0 times higher in the A/PR8/NS124 virus group versus the A/PR8/full NS group. The increased production of the GM-CSF and IFNγ cytokines was observed in both groups when compared to the control group. However, the differences between A/PR8/full NS and A/PR8/NS124 groups in the production of these cytokines were not significant in both cases (p=0.09).
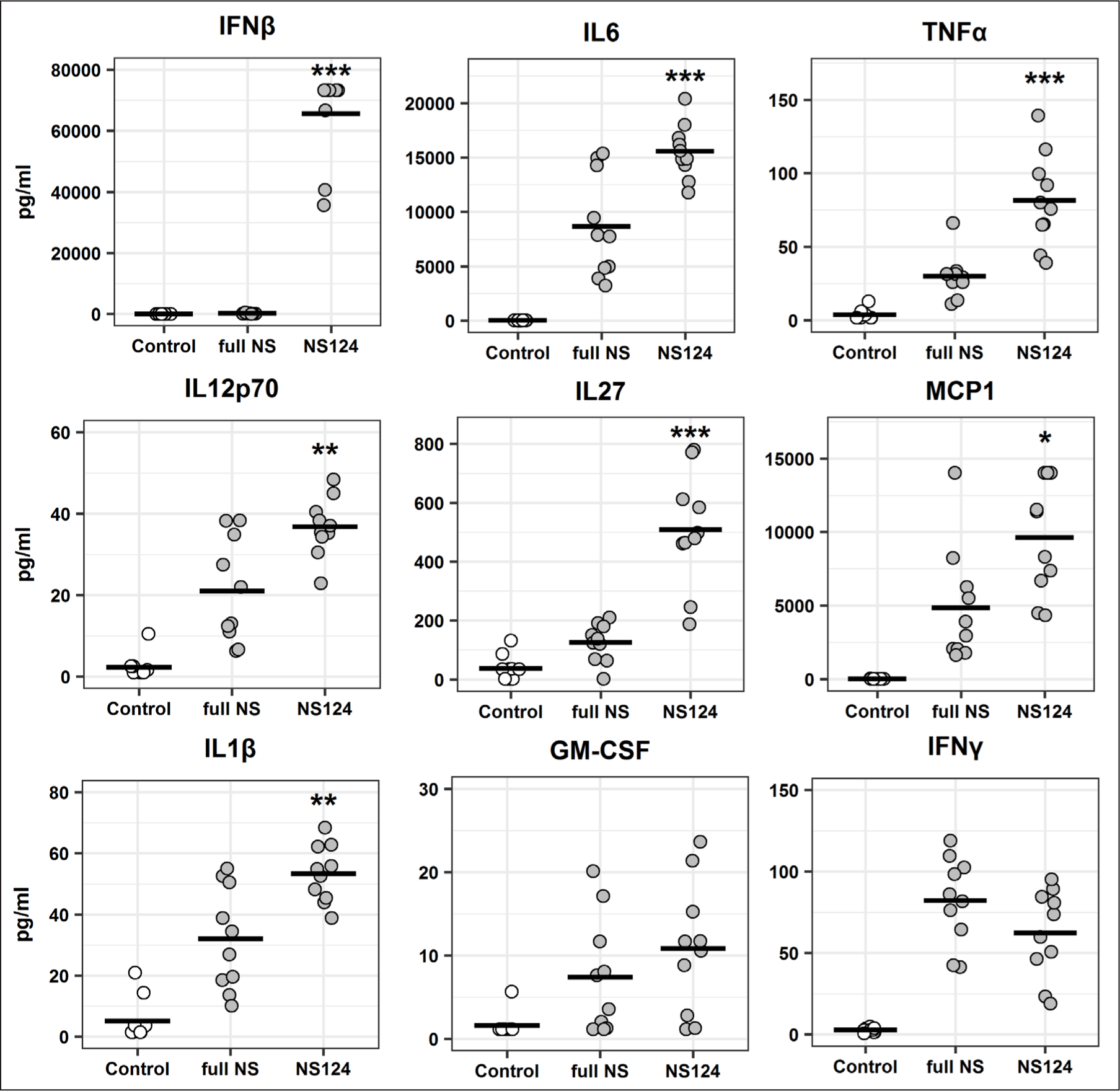
The results are presented as individual values. The horizontal lines indicate the mean values for each group, (n=10). The (*) symbol indicates a significant difference between the full NS and NS124 groups (*: р<0.05, **: p<0.01, ***: p<0.001, Student’s t-test).
The main innate immunity cell populations in peritoneal washings of mice
A panel of markers (CD45, MHCII, CD11c, CD11b, CD24, CD64, and Ly6G) has been used for the identification of the main populations of innate immunity cells in the lungs [18, 19] and other organs (small intestine, heart, kidney, liver, peripheral organs of the immune system) of mice [19]. We used this approach to analyze the cellular composition of peritoneal washings in mice. We have developed a panel of fluorochrome-conjugated antibodies that enable the differentiation of macrophages (CD45+ MHCII+ CD64+ CD11c/CD11b+), dendritic cells (DCs) (CD45+ MHCII+ CD64- CD24+ CD11c/CD11b+), monocytes (CD45+ MHCII- CD64+ CD11c/CD11b+), and neutrophils (SSChi CD45+ Ly6G+) (Fig. 3). Individual subpopulations of DCs were identified by the expression level of markers CD11b and CD103.
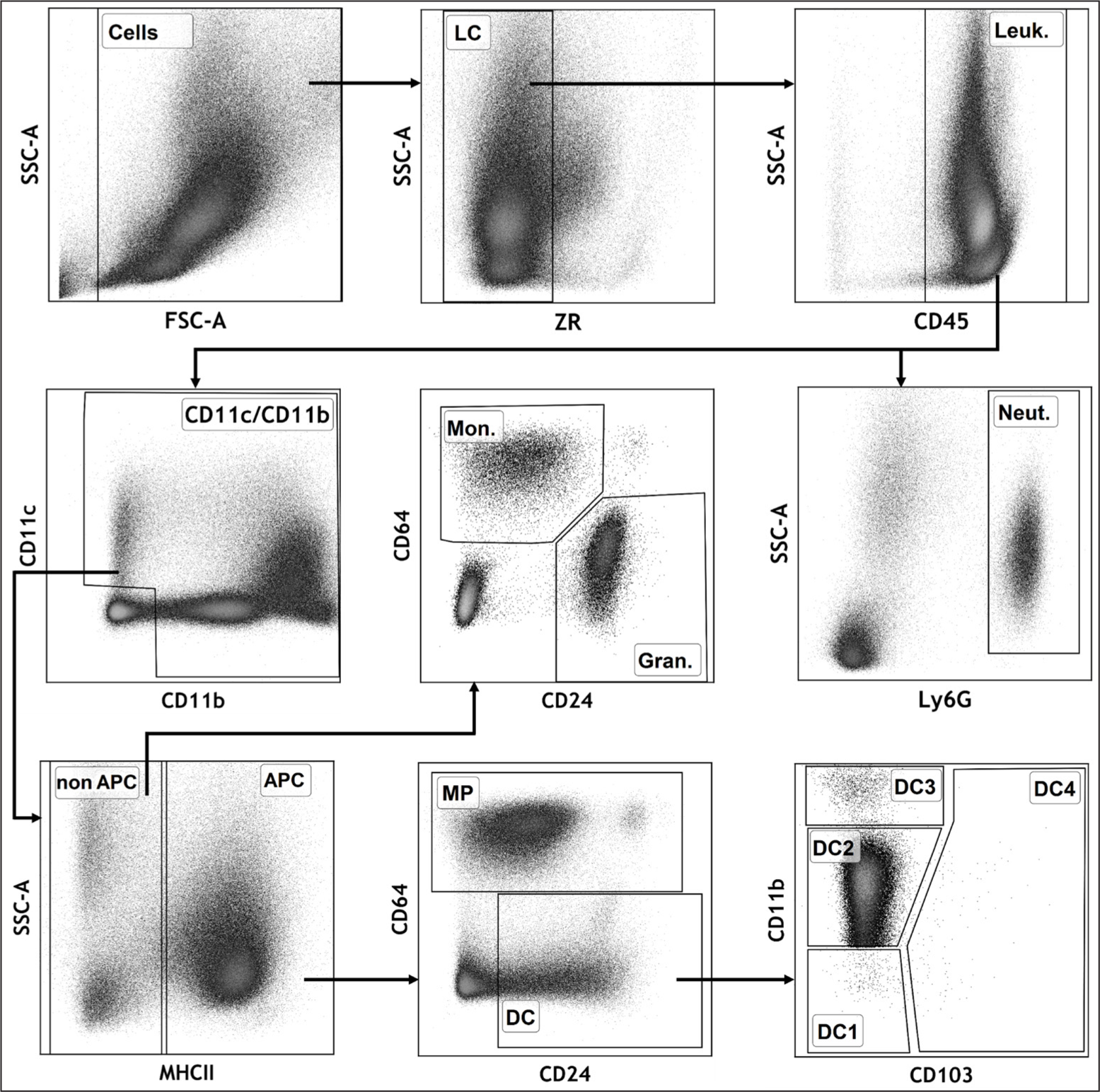
After the elimination of cell doublets and clumps (by FSC-A/FSC-H, not shown) and isolation of the live, single cell population (based on the FSC-A/SSC-A light scattering and binding of the Zombie Red dye), the immunocytes were gated according to the presence of the CD45 marker.
Legend: LC – living cells; Leuk. – leukocytes; APC – antigen-presenting cells (CD45+ CD11c/CD11b+ MHCII+); Mon. – monocytes (CD45+ CD11c/CD11b+ MHCII- CD64+ CD24); Gran. – granulocytes (neutrophils and eosinophils, CD45+ CD11c/CD11b+ MHCII- CD64- CD24+); Neut. – neutrophils (SSChi CD45+ Ly6G+); DC1 – CD11b- DCs (CD45+ CD11c+ CD11b- MHCII+ CD64- CD24+); DC2 – CD11bint DCs (CD45+ CD11c+/-CD11bint MHCII+ CD64- CD24+); DC3 – CD11bhi DCs (CD45+ CD11c+/-CD11bhi MHCII+ CD64- CD24+); DC4 – CD103+ DCs (CD45+ CD11c/CD11b+ CD103+ MHCII+ CD64- CD24+).
Influx dynamics of monocytes and neutrophils in the peritoneal cavity
The relative content of the main populations of innate immunity cells at different time points after the immunization is shown in Fig. 4. Twelve hours after the administration of the A/PR8/full NS, a significant increase was detected in the relative number of monocytes and neutrophils compared with the control group: 2.74±2.48% (0.14±0.10% in control group) and 17.65±11.32% (0.93±0.24% in control group), respectively. In the A/PR8/NS124 immunized group, however, no significant changes in the relative number of these cells were detected. The relative number of macrophages at the same time point after the immunization has significantly changed in both the full NS (p=0.01) and NS124 groups (p=0.05) versus the control group. At 24 h post immunization, a significantly higher relative content of monocytes, macrophages, and neutrophils was detected in both the NS124 and the full NS groups. It should be mentioned that the relative number of the neutrophils at 12 h for the NS124 group was significantly lower than that of the full NS group, whereas at 24 h this tendency remained but the difference was not significant.
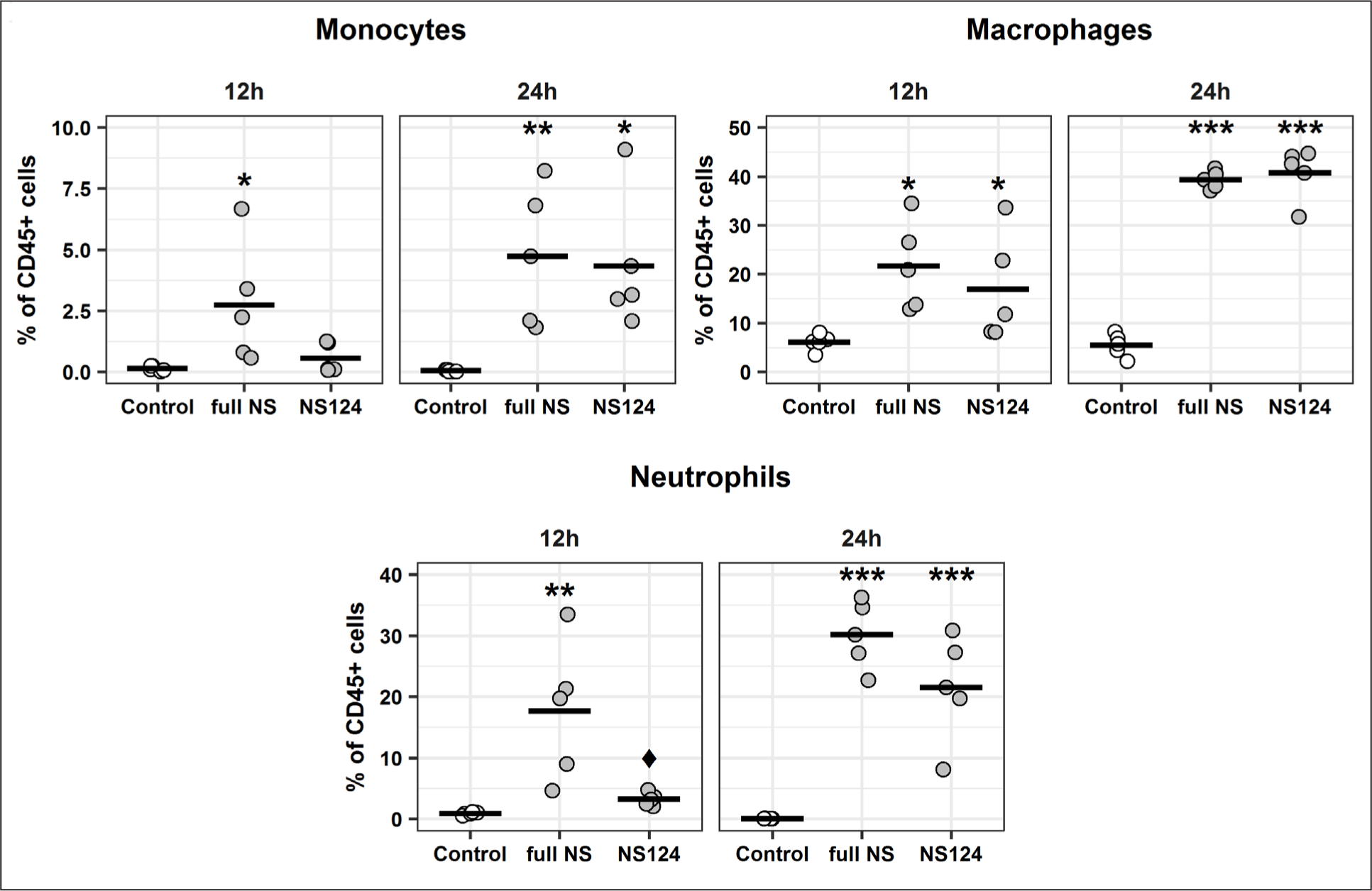
The dots represent the proportions of the corresponding cell populations in individual animals versus the total number of living CD45+ cells. The mean values for the groups (n=5) are shown by a horizontal line. The (*) symbol indicates the significant differences versus the control group (*: p<0.05, **: p<0.01, ***: p<0.001, Dunnett’s test). The (♦) symbol indicates significant differences between the two experimental groups: full NS and NS124 (♦: p<0.05, Student’s t-test).
Relative composition of dendritic cell populations in the peritoneal cavity
In this study, DCs were identified as follows. After the initial gating to retain CD45+CD11c/CD11b+MHCII+CD64-CD24+ cells, CD11b and CD103 markers were used to further identify 4 DC populations (Fig. 3). The DCs, characterized by an intermediate level of CD11b expression (CD11bint DC), made up the largest fraction of DCs in the peritoneal cavity.
Twelve hours after the administration of virus there was a significant reduction in the fraction of CD11bint DCs in the full NS group compared with the control group (p=0.0009) (Fig. 5), while no significant changes were detected in the NS124 group. After 24 h, both groups showed a significant decrease in the CD11bint DCs percentage (p=0.00002, p=0.00003), although in the NS124 group the changes were less pronounced.
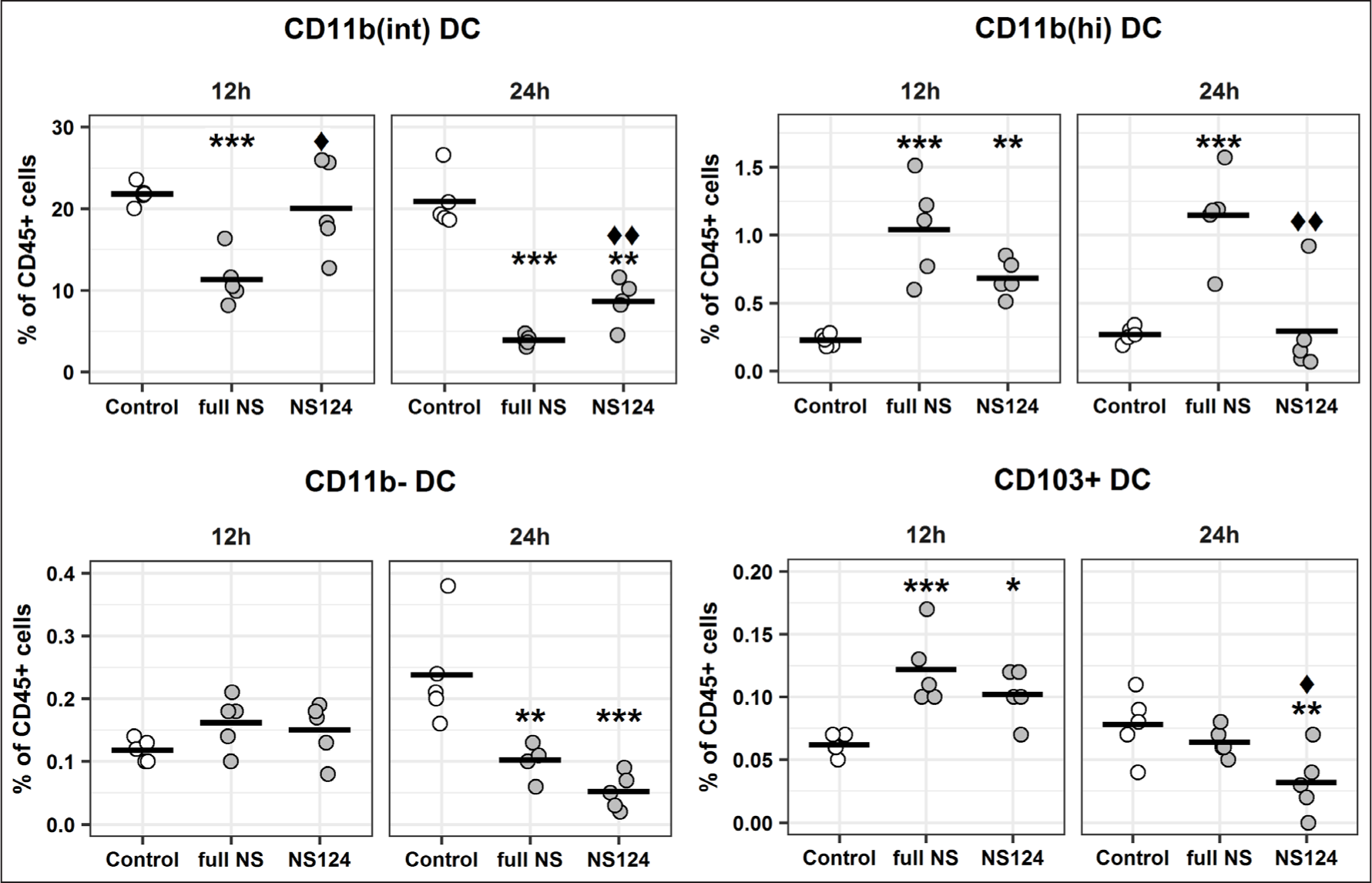
The dots represent the percentage of the corresponding cell populations in individual animals versus the total number of living CD45+ cells. The mean values for the groups (n=5) are shown by a horizontal line. The (*) symbol indicates the presence of significant differences versus the corresponding values for the control group (*: p<0.05, **: p<0.01, ***: p<0.001, Dunnett’s test). The (♦) symbol indicates significant differences between the two experimental groups: full NS and NS124 (♦: p<0.05, ♦♦: p<0.01, Student’s t-test).
The viral administration caused a different response in the minor DC populations, including CD11bhi DCs, CD11bDCs, and CD103+ DCs. An increase in the relative content of these populations occurred 12 h after immunization. In the case of CD11bhi DCs and CD103+ DCs, the differences versus the control group were statistically significant (full NS: p=0.0001, p=0.0007; NS124: p=0.01, p=0.007, respectively). At 24 h post immunization, the percentage of all DC populations in the NS124 group had decreased versus the total number of CD45+ abdominal cells. At the same time, in the full NS group the relative content of CD11bhi DCs remained at a higher level than in the control group and the percentage of CD103+ DCs decreased back to the control group level, while in the NS124 group the CD103+ DC population percentage was significantly lower than in the control group. Thus, 24 h after the immunization with the A/PR8/NS124 strain, a statistically significant decrease in the relative content of CD11bhi DCs and CD103+ DCs was observed (p=0.004 and p=0.02, respectively).
Enhancement of the costimulatory factor CD86 expression
The results of analysis of CD45+ MHCII+ APCs by median fluorescent intensity (MFI) of CD83 and CD86 (APC activation markers), at 12 h and 24 h after immunization, are presented in Fig. 6. It was found that the level of CD83 expression increased in response to the virus administration in both the full NS and the NS124 groups: the statistically significant differences versus the control group (p=0.01, p=0.03) were observed starting from 12 h and lasting up to 24 h after the immunization.
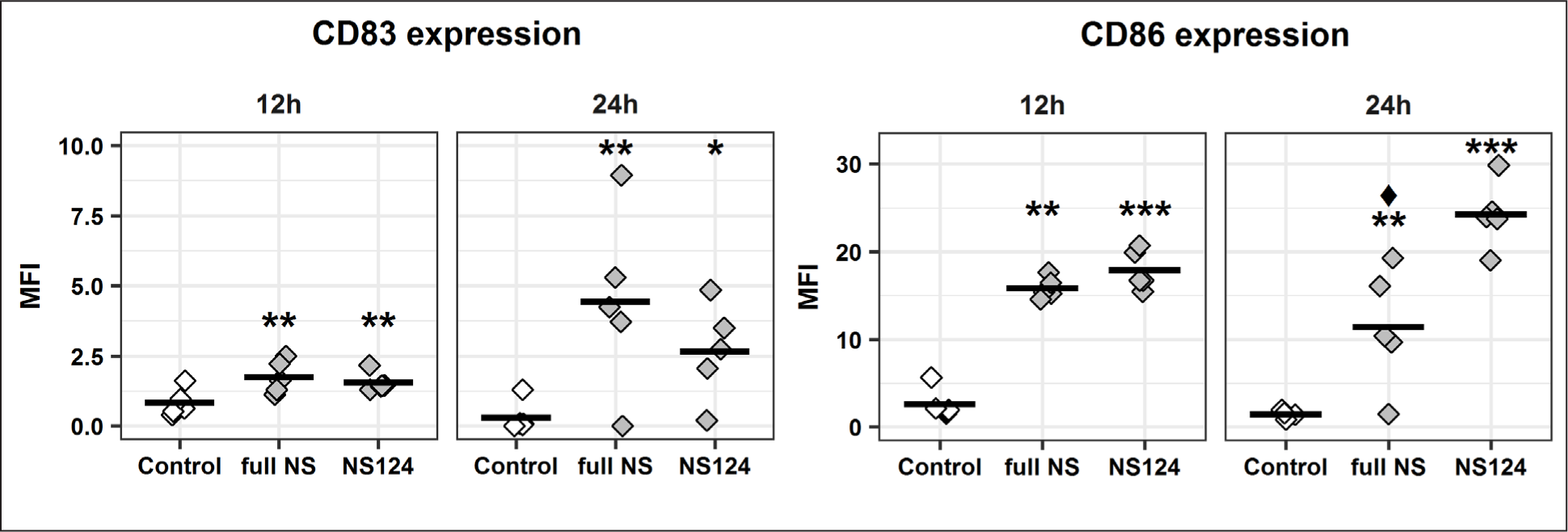
The MFI of the CD83 and CD86 markers for each animal is shown. The mean values for the groups (n=5) are shown by a horizontal line. The (*) symbol indicates the presence of significant differences versus the corresponding values for the control group (*: p<0.05, **: p<0.01, ***: p<0.001, Dunnett’s test). The (♦) symbol indicates significant differences between the two experimental groups: full NS and NS124 (♦: p<0.05, Student’s t-test).
The expression level of CD86 increased considerably 12 h after virus administration. Significant differences versus the control group were observed in both groups of the immunized animals, and the MFIs obtained at this time point had similar values for the full NS and NS124 groups. At 24 h post-immunization, the expression level of CD86 in the NS124 group had increased even more, revealing statistically significant differences not only with the control group (p=0.00003), but also with the full NS group (p=0.006).
Adaptive T-cell immune response
At the final stage of the study, the adaptive T-cell immune responses to the immunodominant epitope of the NP protein were evaluated. It was found that the percentage of antigen-specific CD8+ T-cells in the spleen of mice from the NS124 group was significantly higher than that of the control group and approx. 1.5-2.0 times higher than in the full NS group on the 8th and 21st day after immunization (Fig. 7). At the same time, the biggest differences were due to the changes of the polyfunctional CD8+ T-lymphocytes fractions that were simultaneously expressing IFNγ, IL2, and TNFα cytokines. After 8 days, the results were: full NS 5.9±0.5, NS124 10.7±1.6, p=0.0008; after 21 days: full NS 4.9±0.5, NS124 7.3±0.4, p=0.0003. In addition, on the 8th day after immunization, a higher content of CD8+ T-lymphocyte subpopulations (with the phenotypes IFNγ+ IL2+ TNFα-, and IFNγ+ IL2- TNFα+) was observed in the NS124 group (1.5±0.5 and 8.9±2.6, p=0.05) versus the full NS group (0.5±0.08 and 5.6±1.04, p=0.008). On the 21st day after the immunization, the composition of cytokine-producing cell populations in the NS124 group had changed resulting in an increase of the percentage of IFNγ+ IL2+ TNFα+ and IFNγ+ IL2- TNFα+ T-lymphocytes and a decrease in the relative content of IFNγ+ IL2- TNFαcells (Fig. 7).
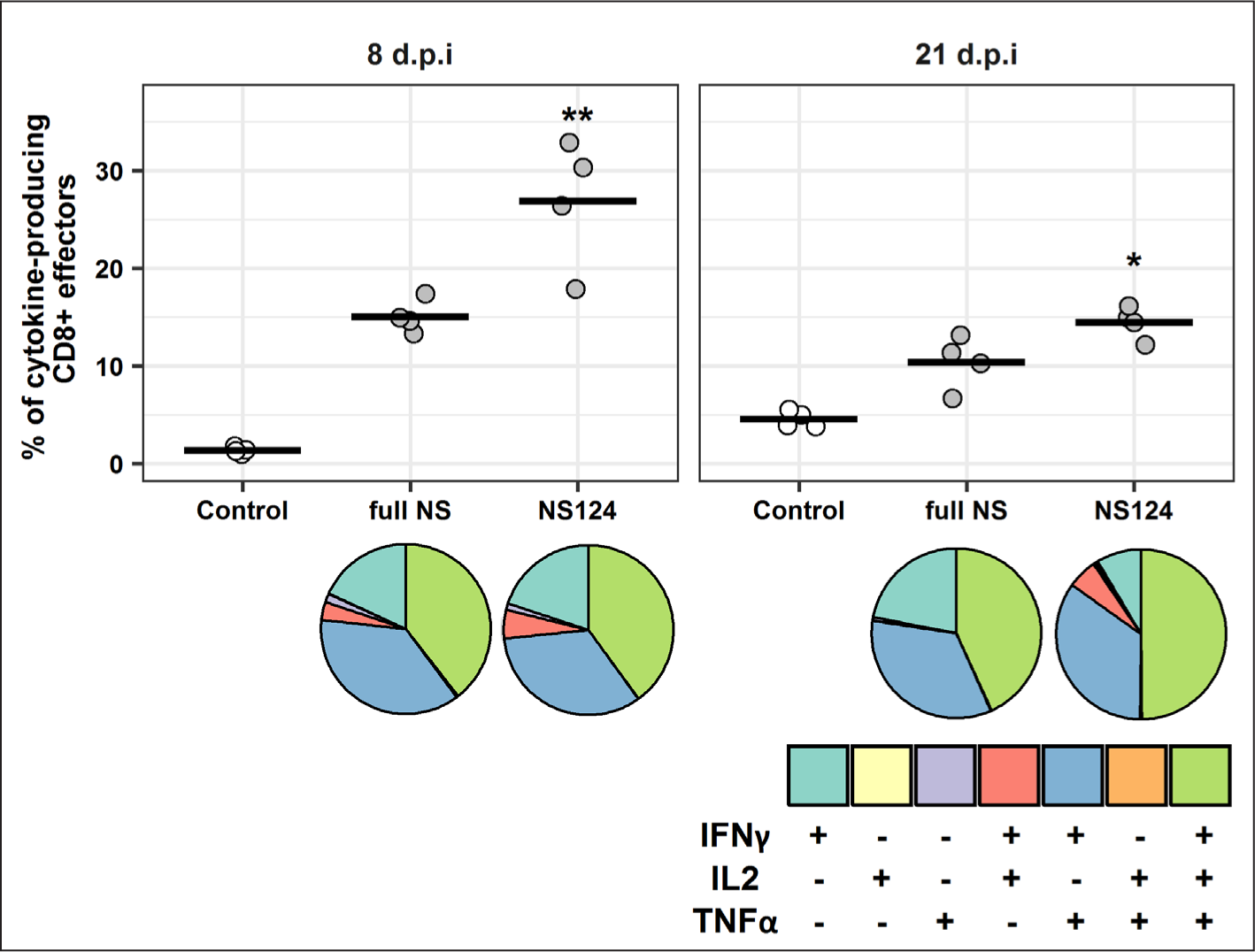
The percentage of the total cytokine-producing T-lymphocytes from the total number of effector CD8+ T-cells (CD8+ CD44+ CD62L-) for the immunized and control groups are presented. The mean values for each group (n=4) are shown by a horizontal line. The relative compositions of populations producing 1, 2, or 3 cytokines simultaneously are shown in the pie charts. The (*) symbol indicates significant differences between the two experimental groups: full NS and NS124 (*: p<0.05, **: p<0.01, Student’s t-test).
DISCUSSION
In the present study, we compared the innate and adaptive T-cell immune responses to influenza A viruses with the full-length NS1 protein and with the NS1 protein shortened to 124 a.a. The comparison of the inflammatory cytokine production in the cells from the peritoneal washings of mice immunized with A/PR8/full NS and A/PR8/NS124 strains showed that shortening of the NS1 protein leads to a decrease of the immunosuppressive activity of the influenza virus. This effect was accompanied by a significant increase in the production of IFNβ, IL6, TNFα, IL12p70, IL27, MCP1, and IL1β in response to immunization with the A/PR8/NS124 strain. The obtained experimental data on the increased production of the IL12 cytokine family members (IL12p70, IL27) is an important contribution to the known information on the high cytokine inducing potential of influenza viruses with the truncated NS1 proteins [4, 20]. The most pronounced difference between A/PR8/full NS and A/PR8/NS124 viruses was associated with IFNβ production, which is suppressed by the interaction of the NS1 protein with the intracellular pattern-recognition receptor RIG-I and other components of type I IFN signaling pathway [21, 22]. Since the production of IFNα/β begins at the earliest stages of the innate antiviral immune response and involves the activation of the expression of a variety of IFN-stimulated genes (ISGs), the increased production of proinflammatory cytokines after immunization with the A/PR8/NS124 virus as observed in our experiments can be probably explained by the high ability of this strain to induce IFN.
Our experiments showed that the intraperitoneal administration of viruses causes the active migration of monocytes, macrophages, and neutrophils into the abdominal cavity. According to the literature data, these cells are involved in the early stage of immune protection against influenza [23]. The resident macrophages accomplish the phagocytosis of infected host cells thereby limiting the spread of the virus [24]. In addition, they attract monocytes that are differentiating into macrophages and monocytic dendritic cells to the nidus of inflammation [25]. Neutrophils also take an active part in the phagocytosis of viral particles and apoptotic cells at the nidus of infection [26]. In addition, they have an important role in attracting cytotoxic T-lymphocytes: according to Lim et al. [27], moving neutrophils leave behind a chemokine trace containing predominantly CXCL12, which directs the migration of T-cells. It is well known that neutrophils perform the MHCI-dependent presentation of viral antigens in mice and also express the CD80 and CD86 costimulatory molecules [28].
Despite the increased induction of cytokines by the A/PR8/NS124 strain, the portion of monocytes, macrophages, and neutrophils in peritoneal washings of mice initially increased faster in response to immunization with the A/PR8/full NS strain. However, by the 21st day after immunization, these parameters had leveled off. It is known that type I IFNs can have both activating and inhibitory effects on the cells of the immune system depending on their concentration and the duration of interaction [29]. In addition to proinflammatory factors, such as the ones measured in the present study, IFNα/β induce the production of a number of anti-inflammatory cytokines as well as the expression of some genes responsible for the induction of apoptosis. It is possible that the hyperproduction of IFNβ, observed after the immunization of mice with A/PR8/NS124 virus, suppressed the migration of neutrophils and monocytes to the peritoneal cavity.
We analyzed the dynamics of the relative composition of the different subpopulations of dendritic cells. These immunocytes are heterogeneous in their composition, origin, location, and function. According to the literature data, over the course of the immune response development, CD11b+ and CD103+ DCs migrate to the secondary organs of the immune system. Experiments that used a recombinant A/PR8 virus containing a green fluorescent protein (GFP) imbedded in the full-length NS1 protein showed that 8.5% of the CD103+ conventional DCs (cDC) of the mediastinal lymph nodes are GFP positive after 48 h post immunization, while among CD11b+ cDCs, only 0.5% of cells contained a fluorescent protein. This may be the evidence of a more intensive migration of CD103+ cDCs to the lymph nodes, although it may also indicate differences in the permissibility of different DC populations for the influenza virus [30].
We used a panel of fluorochrome-conjugated antibodies for the identification of the four subpopulations of dendritic cells based on the expression of CD11b and CD103 markers in our experiments. The analysis of the dynamics in the relative content of CD11bhi DC, CD11bDC, and CD103+ DC populations leads to the suggestion that 24 h after the immunization the DCs leave the viral entry zone. At the same time, a more pronounced decrease in the content of DCs was observed in the NS124 group. That could be an indication of the enhanced migration of DCs to the peripheral lymph nodes for the presentation of the antigen to naïve T-lymphocytes [31, 32].
The analysis of the CD83 and CD86 expression is widely used to study the activation of cells of innate and acquired immunity. It is well known that an increase in the expression of these markers is associated with the APC maturation and the presentation of antigens [33, 34]. We have shown that the level of CD86 expression in mice immunized with the A/PR8/NS124 strain was significantly higher than that in the A/PR8/full NS group at 24 h after the immunization. It is possible that this effect is due to a lack of immunosuppressive activity of the truncated protein NS1, thereby allowing the APCs of animals, immunized with the A/PR8/NS124 virus, to maintain CD86 expression at a high level. On the contrary, in the mice immunized with the strain A/PR8/full NS, the concentration of these markers is reduced at 24 h post immunization.
The ability of the influenza virus containing the truncated NS1 protein to stimulate the higher cytokine production as well as CD86 expression suggests that immunization with A/PR8/NS124 can lead to a more pronounced adaptive CD8+ T-cell immune response to influenza antigens. Our experimental results showed that immunization with A/PR8/NS124 strain induces the formation of a greater number of virus-specific CD8+ T-lymphocytes. The predominant cells among these lymphocytes are the multifunctional CD8+ effector cells (FNγ+IL2+TNFα+ and IFNγ+IL2-TNFα+), which are known for their important role in the formation of the protective antiviral immune response [35].
Thus, based on the obtained data, we conclude that shortening of the NS1 protein leads to an increase in the influenza virus immunogenicity. It contributes to a more effective presentation of viral antigens due to the intensification of dendritic cell migration as well as due to the increase in the CD86 costimulatory factor expression. That, in turn, leads to the formation of a more pronounced adaptive T-cell immune response, accompanied by the formation of polyfunctional effector T-lymphocytes. The results of this study are the theoretical justification for the use of viruses with truncated NS1 protein as live attenuated influenza vaccines and vectors.