
INTRODUCTION
Analysis of the morbidity and mortality patterns of influenza in worldwide practice is based on studies of the age distribution and detection of risk groups [1-6]. At the same time, morbidity and mortality significantly depend on the distinctive genetic characteristics of individual populations and ethnic groups. On the one hand, the direct connection is established between a complicated course of influenza and haplotype HLA (Human Leukocyte Antigenes). On the other hand, the analysis of a number of gene polymorphisms, which determine the level of anti-viral defense, confirms that the contribution of single mutations and single-nucleotide polymorphisms (SNP) to the morbidity and mortality from influenza is significantly higher than was previously considered [7-12]. Large scale studies on population genetics and sensitivity to influenza prove that the anti-epidemic actions in different regions of the country should be planned in accordance to the specific genetic characteristics of population.
It is also evident that in the process of anti-influenza vaccine development it is necessary to consider the possibility of the vaccine’s “genetic” orientation toward large population groups, as well as little to no reaction in response to vaccination in individuals with certain haplotypes of HLA [11,12]. Based on the results of genetic polymorphism analysis it is possible to conclude that in influenza therapy - in the case of mass morbidity, which is distinctive to pandemics - it is necessary to consider the possibility of a complicated course of disease caused by a defect in a certain part of the immune system and/or anti-viral defense. The understanding of genetic basics of pathology of infectious diseases, including influenza, could considerably change the vaccination practice as well as the basics of therapy itself. To that end, it is necessary to systematize the available information on genomes and the corresponding markers, distinctive for the cases with inadequate or pathological reaction to influenza and other accompanying infections. The SNP of the genes, listed and characterized below, is connected with the enhancement of human sensitivity to influenza or complicated course of this disease [9,13,14].
The role of interferon-induced transmembrane protein 3 (IFITM3) gene polymorphism in infectious pathology
Gene (IFITM3). One of the significant discoveries made during the years, passed after the last pandemic, was the detection of polymorphism in gene IFITM3 in groups of patients who developed a severe course of disease, caused by influenza virus A(H1N1)pdm09, with fatal complications in a number of cases (Fig.1) [9, 13-21].

Gene IFITM3 belongs to the family of genes that are induced by type I interferons (IFN). The IFITM3 protein is a transmembrane protein containing two transmembrane domains. Its functional activity is connected with human resistance to the strains of A(H1N1)pdm09 virus and many other infections, including Dengue and West Nile hemorrhagic fevers [13]. The protein IFITM3 exists in different isoforms; one of the most prevalent isoforms lacks the N-terminus domain (Fig.2) [18-21]. Analysis of this phenomenon led to the identification of a splicing site mutation that determines the enhanced human sensitivity to pandemic influenza [19, 20].
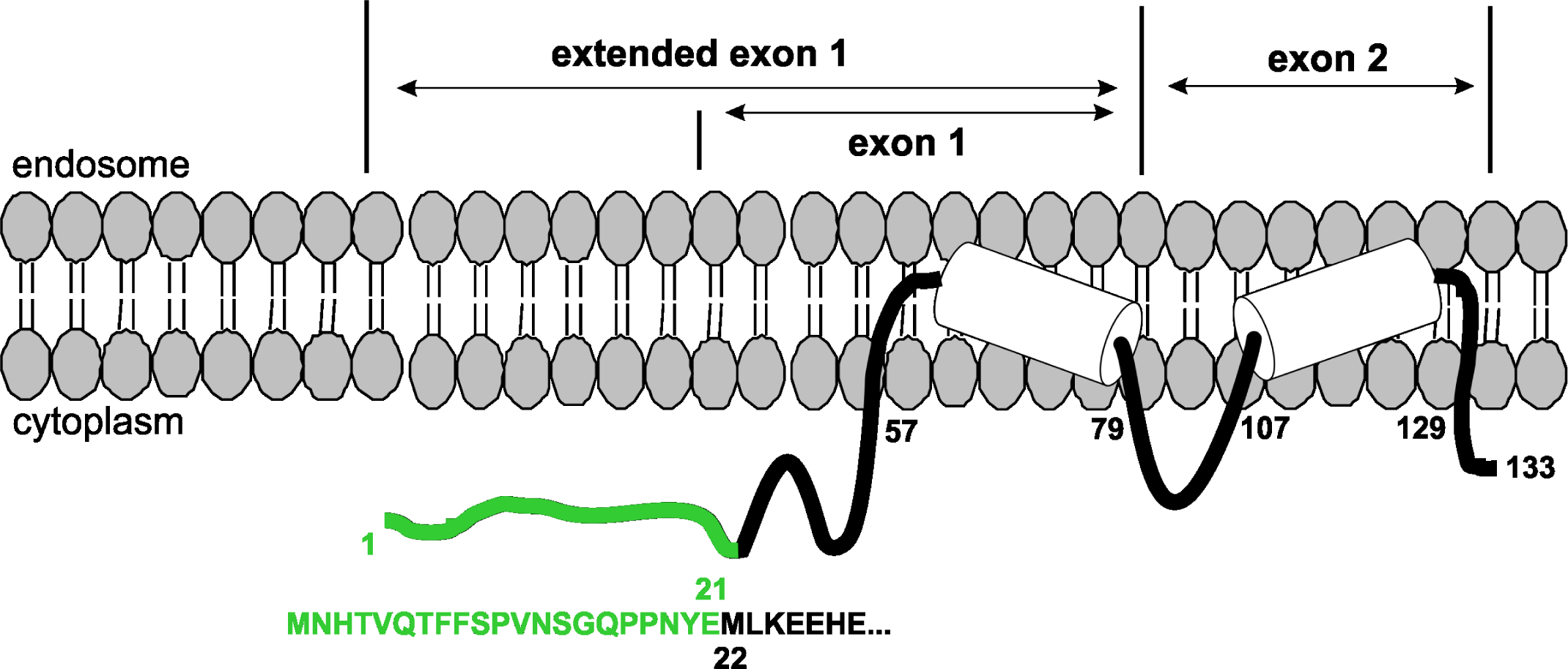
N-end sequence 1-MNHTVQTFFSPVNSGQPPNYE-21, which is absent in the shortened mutant protein, is shown in green.
The analysis of the protein IFITM3 mechanism of action showed that it blocks the cells infection by preventing the entry of viral particles via endocytosis. [22]. This protein was shown to suppress the infection of Ebola, HIV-1 (human immunodeficiency virus type I), hepatitis C, and Dengue viruses [13]. It was also shown that IFITM3 protein suppresses the S-protein dependent endocytosis of Middle East Respiratory Syndrome coronavirus (MERS-CoV) [23], preventing the virus genetic material from entering the cell.
The wide spectrum of IFITM3 protein antiviral activity is caused by the protein’s profound effect on the stability of complex ATPhase of vacuoles (v-ATPhase) and endosomes. The interaction of v-ATPhase with endosomes, which leads to the relocation of Clatrin and pH reduction, plays an important role in the endocytosis of viruses [24]. This process is extremely attractive as a drug discovery target because it plays a key role in the cell infection process. It was established that classical Clatrin and v-ATPhase inhibitors are effective viral reproduction inhibitors and belong to broad-spectrum class of medicines. It turned out that Arbidol also belongs to this group of inhibitors [25]. Thus, one more mechanism of Arbidol action was revealed, which could explain its wide spectrum antiviral activity. However, the interaction of protein IFITM3 with viruses on the Clatrin pathway of virus particles internalization on the early infection phases could not be restricted to the mechanism of antiviral defense.
Two different transcripts could be formed during the expression of the IFITM3 gene – the full transcript and the shortened transcript version, which encodes the protein sequence, lacking the N-end 21 amino acid fragment [21].
In general, the IFITM3 protein is a restriction factor for virus reproduction, acting by the formation of the cells resistance to the different types of viruses. However, the detailed mechanisms of anti-viral defense formation in cells remain unknown in spite of the protein IFITM3 discovery. As it was shown in a number of publications [18-20], mice with silenced IFITM3 gene had a more complicated course of influenza than mice with wild IFTIM3 gene. There is a known mutation of this gene in humans: substitution of Thymidine by Cytosine in the 1st intron splicing site [18].
The connection of this mutation to the severity of influenza A(H1N1)pdm09 was studied in a European population group [19]. In this study, the frequency of hospitalizations was the criteria of complications during the development of infection. Patients who were hospitalized with severe influenza and/or the complications caused by influenza were found to have an enhanced frequency of homozygosity for a rare C allele of gene IFITM3. The frequency of SNP rs12252-C in patients with severe influenza totals 5.3% versus 0.3% (which is common) for the European population. Interestingly, the frequency of occurrence of SNP rs12252-C in Chinese populations was significantly higher. The genotype frequency CC reached approx. 25% in the Chinese populations. Among the patients in China who had severe forms of influenza the occurrence of genotype CC reached up to 69% (Fig.3).
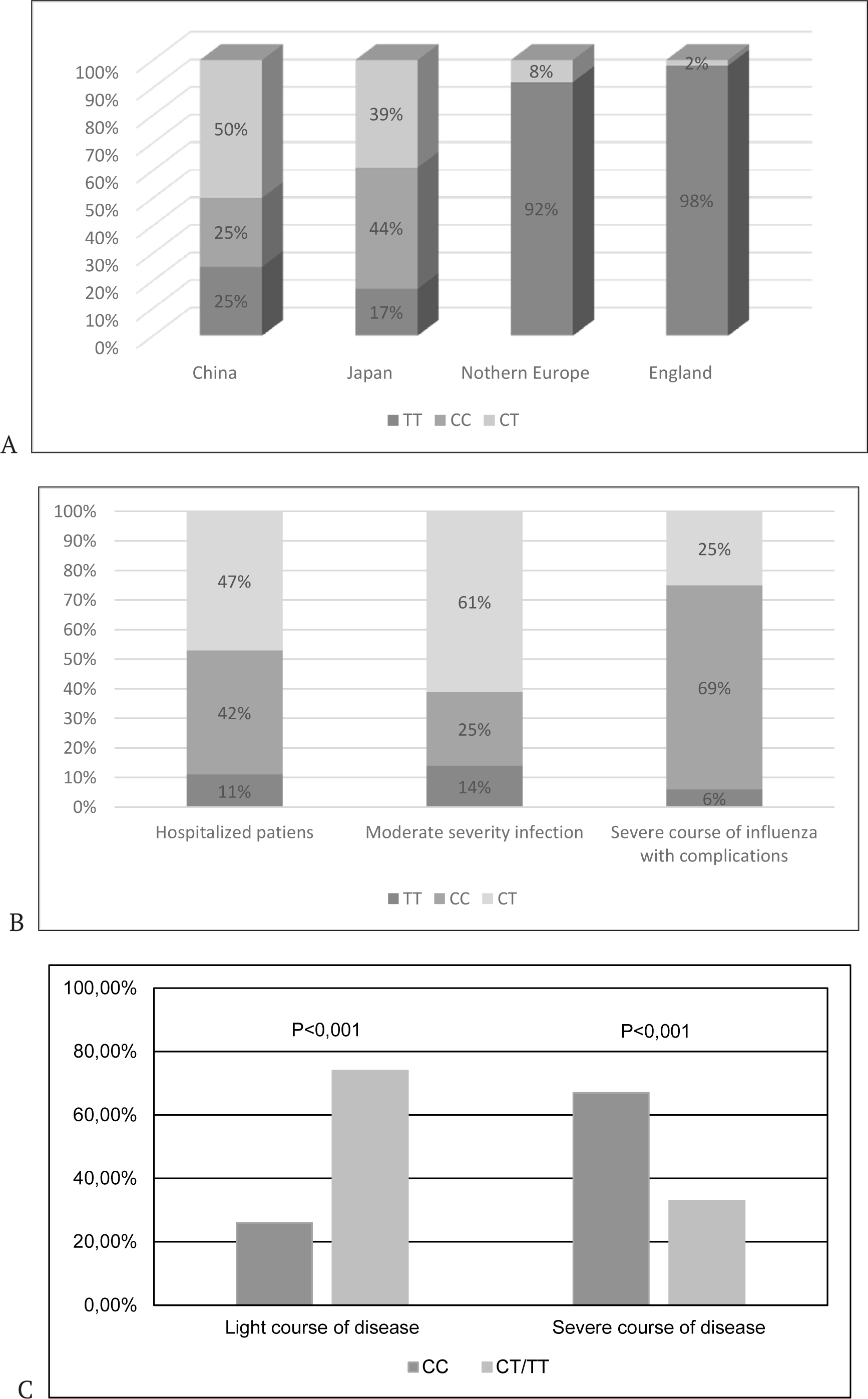
B - groups of patients: 1 - hospitalized patients, 2 - moderate severity influenza, 3 - severe course of influenza with complications;
C - the frequency of alleles occurrence in patients with A(H1N1)pdm09 virus.
The results on allelic frequencies of the IFITM3 gene described here were obtained by the authors by the statistical data processing of 1,000 genomes [19]. The IFITM3 allelic frequency varied significantly in different populations. However, among members of the Han population, originating in the Southern province and inhabiting Eastern, Southern, and central China, the frequency of CC genotype rs12252 has been found to be as high as 69%. As a result of the detailed analysis of IFITM3 allelic frequency in the population and influenza morbidity (mortality), the authors come to the conclusion that the high frequency of occurrence of CC genotype contributes to the epidemiology of influenza in China. As a matter of fact, it is precisely in China that high frequencies of massive influenza breakouts are recorded that often lead to pandemics.
The discovery of correlation between genotype IFITM3 rs12252 and the clinical pattern of influenza infection prompted interest to study the separate components of “cytokine storm” in patients with the variant IFITM3 gene [21]. These studies were conducted with patients infected with the pathogenic strain of A(H7N9) influenza virus that is currently in circulation. The patterns of expression of the following markers of “cytokine storm” were analyzed: MPC-1 (Monocyte Chemoattractant Protein-1), IL-1β (Interleukin-1β), IL-6, IL-8, IL-10, TNF-α (Tumor Necrosis Factor), IFN-γ and C-reactive protein [27]. The authors studied the level of content of these proteins in peripheral blood. As a result of this study, it was established that the patients with genotype CC have more active synthesis and higher level of the secretion of protein MPC-1 in comparison with patients with CT or TT genotypes (Fig.3). The severity of disease also had direct correlation with these parameters – genotype CC and excessive nonspecific immune response. Furthermore, the authors have conducted the study of cytokines (MIP-1α, MIP-1β, IL-1β, IL-6 and IL-8) in the lungs of dead patients. It turned out that the level of content of some pro-inflammatory cytokines in lungs was 100–1,000 times higher than in the peripheral blood. Thus, the development of “cytokine storm” is aided by IFITM3 rs12252 gene mutation.
To conclude the discussion of the role of gene IFITM3 polymorphism in infectious pathology it should be mentioned that the SNP rs12252 contributes to the development of Kawasaki syndrome – the early children’s vasculitis [15].This abnormality is also common for certain population groups of South-East Asia and it could lead to a severe complication, such as aortic aneurysm [28]. It should be mentioned that influenza infection provokes vasculitis, including cerebral vasculitis, which suggests that the influenza infection affects the development of the pathology of the cardiovascular system.
Polymorphism of genes, which make an additional contribution to the severity of influenza infection
Gene polymorphisms have been studied in order to understand the susceptibility of populations to infectious diseases and their severe progression. These studies have fundamental significance for pediatric practice and global epidemic processes [9,14, 17]. The group of genes that have an impact on the development of complications over the course of influenza, is significantly widening over the years [9,17]. Table 1 presents a list of these genes and their probable role in the disruption of certain functions contributing to the eradication of influenza and other viral infections.
| Gene | Functions of the encoded protein | Defect | Reference |
|---|---|---|---|
| IFITM3 | Factor of antiviral defense on the endosomes level | Defect of inner cell’s antiviral defense on the initial infection step (endosomes) | [18,19,21, 25] |
| SP-B | Surfactant protein B – lungs surfactant | Pneumocytes defense, stability of teeth ridge, oxygen transport and clearance of viruses and bacteria | [29,30] |
| FCGR2A | Fc-receptor – factor of infectious virus clearance | Defect of virus clearance | [31,32] |
| C1QBP | Factor of the complement system | Defect of the complement system and complement dependent cytolysis of infected cells | [8,9,33] |
| DAF/CD55 | Factor of the complement decom-position acceleration, antigen of the Kromer blood group system | Defect of the natural mechanisms of the lungs’ defense from the damage caused by complement in the course of influenza infection | [34] |
| MBL2 | Mannose-binding cell lectin | Natural immunity regulation defect | [35] |
| SOCS4 | Suppressor of the cytokine dependent signal systems | Defect of control of cytokine synthesis and activity (possible development of the “cytokine storm”) | [27,36] |
| SECISBP2 | Complex of Se – dependent enzymes | Defect of the antioxidant system | [37] |
The lung surfactant proteins play an important role not only in providing the stability of oxygen transport but also in antibacterial and antiviral defense. Long-term clinical observations showed that polymorphisms in genes that encode the surfactant proteins, and in particular the protein B (SP-B), play the key role in susceptibility to infections like influenza [29]. It was also established that the polymorphism of the SP-B gene is connected with a severe course of infection caused by the respiratory syncytial virus [30].
The role of complement system factors in bacterial and viral infections is very well known. In this respect, the role of the C1QBP gene polymorphism (Complement Component 1, Q subcomponent Binding Protein) in the complications of the course of influenza [8,33] is worth close attention. The gene C1QBP is encoding the protein C1QBP, which is a homotrimer in mature form. The structural particularity of homotrimer C1QBP lies in asymmetric charge distribution on the surface of the molecule. This protein is interacting with a wide range of molecules involved in regulation of immune system: CDK13 [38], HRK [39], VTN [40], NFYB [41], FOXC1 [42], DDX21, DDX50, NCL [43], SRSF1, SRSF9 [44], CDKN2A [45] and other proteins including CD93 [46]. The function of C1QBP consists in antibody dependent cytolysis of infected cells with the involvement of complement and activation of phagocytosis. This protein is accumulated in mitochondrions during viral infection and inhibits signal transmission, which depends on RLR (RIG-I-like receptors), by interaction with the antiviral protein MAVS (Mitochondrial Anti-Viral signaling protein). The protein C1QBP is involved in the activation of blood coagulation cascade [47]. The action of C1QDP has pleiotropic character and depends on infection stage.
CD55 (Complement Decay-Accelerating Factor; DAF) – acceleration factor for the complement decomposition – antigen of the Kromer system of blood types. DAF/CD55 carries out anti-inflammatory functions by protecting cells from damage by a complement system as well as by the control of leucocytes migration to the inflammation center [47]. DAF/CD55 is expressed in vascular endothelium, mononuclears of peripheral blood and also in epithelial cells (including lung and endometrial epithelia) [48]. CD55 inhibits activation of C3 and C5 components of complement system [49]. Significant level of expression of CD55 in respiratory epithelium stresses the importance of lungs protection from the damage caused as a result of excessive activation of the complement system. It is notable that the expression level of CD55 is regulated by progesterone and estrogen [50].
SNP in the gene’s CD55 (rs2564978 genotype T/T) promoter is associated with more severe course of influenza caused by A(H1N1)pdm09 virus [34]. As was established by experiments in vitro, the infection of cells by the influenza virus A(H1N1)pdm09 causes the intensification of the protein CD55 expression. Patients with T/T rs2564978 genotype contain much less of CD55 on the surface of mononuclears of peripheral blood as compared with the patients with C/C and C/T genotypes [51].
The activation of the complement system makes a significant contribution to the lung tissue damage over the course of influenza infection. High levels of C3, C5b-9 and MLB were detected in the lungs of mice infected with influenza A(H5N1) viruses, while the administration of C3aR antagonist significantly reduced the degree of inflammation in the lung tissue [52].
The mutation in the gene’s CD55 promoter, which apparently leads to the reduction of its expression, disrupts innate lungs defense mechanisms against the damage induced by complement during influenza infection leading to a higher chance of severe disease course and lethal outcome.
Discussion of the correlation of human genes polymorphism with severity of influenza course brings up the problem of inadequate reaction of the innate immunity to infection. It is known that the activation of transcription of genes, encoding the proinflammatory cytokines, plays an important role in pathogenesis of severe influenza [21].
The system of genetic control of the innate immune response towards influenza infection as well as the functions of genes that are involved in downward regulation of the expression of genes, encoding the proinflammatory cytokines is of particular interest.
Gene SOCS4 (Suppressor of Cytokine Signaling 4) holds one of the key positions in the hierarchy of genetic control of the activation of proinflammatory cytokines synthesis [36].
The interest in the genetic control of these processes is natural. Recently, it was established that the genes of SOCS family play an important role in restraint of nonspecific natural immune response to different pathogens. Particularly, it primarily relates to the protein SOCS4 – the suppressor of cytokine signal systems 4. As a downward regulator of cytokine synthesis control, this protein belongs to the key factors of defense from excessive proinflammatory reaction to influenza virus. The proteins of this family control the innate and adaptive immune response by inhibiting the signaling pathway JAK/STAT [36].
SECISBP2 – gene, encoding the enzymatic complex (Sec Insertion Sequence Binding Protein 2), which in turn ensures the inclusion of selenocysteine in the protein structure [37]. Enzymes containing selenocysteine are the members of the cell’s Red-Ox defense system from oxygen radicals. Generation of oxygen radicals is the cells reaction to the majority of pathogenic microorganisms including viruses [37]. In the case of hereditary mutations in SECISBP2 gene, the acceleration of oxygen active forms synthesis leads to the systematic enhancement of cellular sensitivity to insulin. This is observed in mice with knocked out gene that encodes the antioxidant enzyme glutathione peroxidase 1, which contains selenium. The presence of gene SECISBP2 mutations in humans is clinically manifested by azoospermia, axial muscular dystrophy, disorder of T-lymphocytes proliferation and suppression of immune system in general. A high degree of lipid oxidation by peroxide and DNA oxidative damage, reduction of DNA damage reparation potential and shortening of telomeres are observed in individuals with these mutations. The pleiotropic effect of mutations in SECISBP2 gene is caused by the disruption of functions of the whole selenoproteome [37]. This hereditary defect should be especially prominent in the case of influenza infection and influenza-caused pneumonia. It is known that the influenza antioxidant therapy can provide significant clinical results in the area of intoxication syndrome relief and prevention of cardiovascular complications [52].
CONCLUSION
The understanding of genetic polymorphism in certain genes enables to direct the development of the systematic influenza therapy with the focus on phenotypic changes, caused by mutations, with consideration of the distribution degree of the hereditary susceptibility to severe course of influenza among groups of genetic risk and population, in general.
In spite of the possibility of the wide distribution of mutations in the identified group of genes, related to the enhanced sensitivity to influenza, it should be recognized that the presentation level of the viral antigens within the certain population groups with the different types of HLA [10, 12] remains most important.
Mortality from influenza is registered in the acute period of the illness and usually on the peak of the epidemic, and then, as a “postponed mortality”, in the period from 1 to 3 months after the course of the disease. Most commonly, the lethal outcome is connected with the accompanying diseases, usually with cardiovascular pathology - namely heart attack and stroke.
Thus, these factors should be taken in account while analyzing the genetic background of the population.