
Introduction
The beef industry is of significant value to the Irish economy, with annual beef exports worth in excess of €2.1 billion (BordBia, 2022). The Irish beef industry is unique in that over 60% of beef cattle slaughtered in Ireland originate from dairy herds, while the remainder comes from suckler beef-based systems. In contrast, a much lower percentage of red meat originates from dairy herds in continental Europe (Maher et al., 2021).
Currently, there are four national selection indices for cattle: (1) Economic Breeding Index (EBI), which is used to select parents to breed profitable animals for dairy herds (Berry et al., 2007); (2) Dairy-Beef Index (DBI), which is used to select superior beef sires to use on dairy cows, thereby improving profitability for dairy and beef herds (Berry et al., 2020); (3) Terminal Index, which is designed to identify the most profitable sires for mating on beef cows whose progeny are intended for slaughter (Kelly et al., 2020); and (4) Replacement Index, which is designed to identify the most profitable parents of animals destined to become dams for beef herds (McCabe et al., 2020). All four national selection indices include carcass traits (i.e. carcass weight, conformation and fat score), but with varying emphasis.
Animal breeding and genetic selection have had both favourable and unfavourable impacts on a range of traits and species. Positive impacts can be seen in the genetic gain observed in milk production in dairy cows (Berry et al., 2016), and in carcass traits in broilers (Tavárez & Solis de los Santos, 2016). Nevertheless, deterioration in health and fertility traits in dairy cows has been observed as a result of selecting for traits antagonistically related to them (Pritchard et al., 2013). For example, breeding has had an unfavourable impact on cow fertility in the US Holstein (HO) herd due to intensive selection for improving milk traits (Lucy, 2001). Through the evaluation of fertility trends and greater emphasis on its importance, multi-trait selection indices have helped to overcome the issue of declining fertility in dairy cows (Berry et al., 2014). As a result, improvements in genetic gain have since been observed for fertility in cattle (Berry et al., 2016), highlighting the importance of breeding programmes and improved genetic selection. With respect to carcass traits, Twomey et al. (2020a) documented favourable genetic trends in Irish beef cattle originating from beef herds, but a decline in carcass traits in animals originating from dairy herds. Different trends in carcass traits were observed in different beef populations due to varying selection pressures in breeding programmes for dairy and beef animals.
Breeding in animal populations is both cumulative and permanent, with long-term benefits, as well as unintended consequences on antagonistically related traits. Berry (2018) has shown that breeding can account for half the genetic gain or genetic loss in livestock performance. The improvements, deterioration or unintended consequences directly associated with animal breeding have been shown to have a direct impact on farm productivity and profitability (Nielsen et al., 2013). To our knowledge, this is the first study to assess both the genetic and phenotypic trends of carcass traits within gender of animals originating from dairy and beef herds. In addition, it is unknown whether genetic improvements, or deterioration, is due to a breed shift in these animals. Therefore to determine the effectiveness of current Irish breeding programmes, the objective of the present study was to estimate the genetic and phenotypic trends in carcass traits in Irish cattle within dairy, dairy-beef and suckler beef animals, as well as assess the impact of a breed shift on the genetic trend.
Materials and methods
Data
Ancestry and breed composition was obtained for 22,948,551 animals born between 1986 and 2020 inclusive, from the national database maintained on behalf of the industry by the Irish Cattle Breeding Federation (ICBF).
Estimated breeding values
Estimated breeding values (EBVs) for carcass traits were regenerated using phenotypic and pedigree data from the September 2020 Irish carcass genetic evaluations in Mix99 (Strandén & Lidauer, 1999), which included phenotypic data on 20,236,674 animals. The carcass evaluation consists of three multi-trait animal models. Firstly, the goal traits of carcass weight, cow live weight and cull cow carcass weight were generated using a 12 linear animal model which also included 150–250 d weight, 250–350 d weight, 350–450 d weight, 450–550 d weight and 550–700 d weight, skeletal score, foreign weaning weight EBV, foreign skeletal EBV and foreign carcass weight EBV as predictor traits. Secondly, the goal trait carcass conformation was generated using a nine-trait linear animal model, which also included muscle score, calf quality, calf price, weanling price, post-weanling price, foreign muscle EBV and foreign skeletal EBV as predictor traits. Thirdly, the goal trait carcass fat was generated using an 11 trait linear animal model which also included feed intake, carcass weight, carcass conformation, carcass fat, 350–450 d weight, 450–550 d weight, 550–700 d weight, skeletal score, foreign weaning weight EBV, foreign carcass weight EBV and foreign carcass conformation EBV as predictor traits. Animals with an unknown sire or dam were not considered further; 14,570,772 animals remained.
Phenotypic data
Carcass data, including carcass weight, conformation score and fat score, were available for 22,954,764 cattle slaughtered between the years 2002 and 2020, inclusive. Carcass records were divided into two categories: (1) prime, animals that had never parented an animal, and (2) cows, females with at least one recorded calving event. For prime animals, only heifers and steers slaughtered between 12 and 36 mo of age, inclusive, and young bulls slaughtered between 12 and 24 mo of age, inclusive, were retained. Carcass records for cows were those from females slaughtered between 24 and 180 mo of age with at least one calving record. Animals with a carcass weight of <100 or >800 kg were discarded. Only animals that had an EBV for carcass traits as well a known sire and dam were retained. The final dataset after edits, consisted of 3,528,392 steers, 2,341,262 heifers, 1,154,377 young bulls and 1,784,647 cows.
Genotype definition
An animal’s genotype was defined based on the breed proportions of their sire and dam. Dams and sires were defined as dairy if their breed composition was ≥87.5% of a dairy breed (i.e. HO, Friesian [FR], Jersey [JE], Ayrshire [AY], Norwegian Red [NR], Montbeliarde [MO], Brown Swiss [BS], Normande [NO] and Rotbunt [RB]). Dams were defined as beef if their breed composition was ≥45% of a beef breed (i.e. Aberdeen Angus [AA], Belgium Blue [BB], Charolais [CH], Hereford [HE], Limousin [LM], Simmental [SI], Aubrac [AU], Blonde d’Aquitaine [BA], Piedmontese [PI], Parthenaise [PT] and Saler [SA]). Sires were defined as beef if their breed composition was ≥87.5% of a beef breed. Animals with either a dam or a sire not defined as either beef or dairy were not considered further. Animal genotypes were classified into three categories: (1) dairy breed animals (i.e. dairy dam and dairy sire [D×D]), (2) dairy-beef animals (i.e. dairy dam and beef sire [D×B]) and (3) suckler beef animals (i.e. beef dam and beef sire [B×B]; Ring et al., 2018).
Data analysis
Genetic trends
The mean annual EBV for carcass weight, carcass conformation, carcass fat, cow live weight, cull cow carcass weight and cull cow carcass conformation for each animal was estimated based on the year of birth of the animal. To ensure an adequate representation of animals per year; animals born prior to 1990 or after 2018 were not considered further for D×D animals (n = 4,733,447), animals born before 2002 or after 2018 were not considered further for D×B animals (n = 1,963,903). Finally, animals born prior to 1994 or after 2018 were not considered further for B×B animals (n = 7,113,683). A linear regression model was then used to separately estimate the linear genetic trend for these animals from 2008 to 2018 for each of the three different breed genotypes. The dependent variables were EBVs for carcass weight, carcass conformation, carcass fat, cow live weight, cull cow carcass weight and cull cow carcass conformation; birth year of the animal was included as a fixed effect. Breed substitution refers to replacing one breed in a population with another breed (Haskell et al., 2014). To investigate the effect of breed substitution, breed was included as a fixed effect, which included the percentage fitted as a covariate (including breeds AA, AU, BA, BB, CH, HE, LM, PI, PT, SA, SI, AY, BS, HO, FR, JE, MO, NO, NR and RB). For the purposes of the present study, genetic change was attributed to breed substitution if genetic change was different when breed was included as a fixed effect compared to when it was not.
Phenotypic trends
A linear regression model was used to estimate phenotypic trends for young bull, heifer and steer slaughter records for carcass weight, carcass conformation, carcass fat and age at slaughter; year of birth and age in months were included as fixed class effects. Least square means were estimated for each year of birth for each of the carcass traits. For carcass weight, conformation and fat score, young bulls were adjusted to 18 mo of age, heifers were adjusted to 24 mo of age and steers were adjusted to 28 mo of age. For age at slaughter, the linear regression model included carcass weight instead of age in months; least square means were estimated for the average age per year for the average carcass weight of each gender within each genotype (Table 1). Using a linear regression model phenotypic trends were also estimated for cow slaughter records for carcass weight, carcass conformation and carcass fat. Year of birth and parity were included as fixed effects. Yearly averages for each of the carcass traits for cows were estimated using least square means when parity three was the reference animal. To estimate the linear phenotypic trend for each trait for birth years in a 10-yr period, year was included as a continuous fixed effect in a separate analysis within each of the three genotypes (D×D), (D×B) and (B×B) for steers, young bulls, heifers and cows.
Mean carcass weight by genotype used in calculating least square means for age at slaughter
| Carcass weight (kg) | |||
|---|---|---|---|
| Genotype | Steers | Young bulls | Heifers |
| Dairy dam and dairy sire | 330 | 290 | |
| Dairy dam and beef sire | 340 | 340 | 280 |
| Beef dam and beef sire | 380 | 400 | 330 |
Genetic trends using animals with phenotypic records were also estimated. A linear regression model was also used to estimate the trends in EBVs for carcass weight, carcass conformation and carcass fat for animals with a phenotypic record for young bulls, heifers and steers, separately. Similarly, the genetic trend for cull cow carcass weight and cull cow carcass conformation was also estimated for animals with a phenotype record.
Results
Breed proportions for each of the three genotypes are presented in Appendix Figure A1. Approximately 90% of the breed composition for D×D animals was represented by HO and FR. The proportion of HO and FR has fluctuated over the last 28 yr. The percentage of HO reached its peak in the early 2000s but has since decreased by approximately 10%. In relation to D×B animals, there are five main beef breeds, as well as the proportion of dairy. Usage of LM, BB and CH combined has decreased by 16% in the dairy herd since 2002, while both AA and HE combined saw an increase in usage of 17%. Finally, since the year 2000, there has been an increase in the proportion of LM (10%) and a decline in CH (10%) breed composition for B×B animals. There has also been an increase in the percentage of dairy breed composition for B×B animals.
Genetic trends
The genetic trend for carcass traits in the national cattle population is presented in Figure 1. The trend for carcass weight showed a decrease of 0.47 kg per year for D×D animals between the years 2008 and 2018. When breed covariates were included in the model, the D×D genotype had a similar genetic trend for carcass weight, with an observed decrease of 0.34 kg per year. Similarly, the genetic trend for D×B animals for carcass weight showed a decrease of 0.14 kg per year since 2008. However, when breed was accounted for, the genetic trend for D×B animals changed from 0.14 kg to 0.31 kg per year. The B×B animals carcass weight showed an increase of 1.1 and 0.84 kg per year when breed covariates were not included and included in the analysis, respectively, from 2008 to 2018.
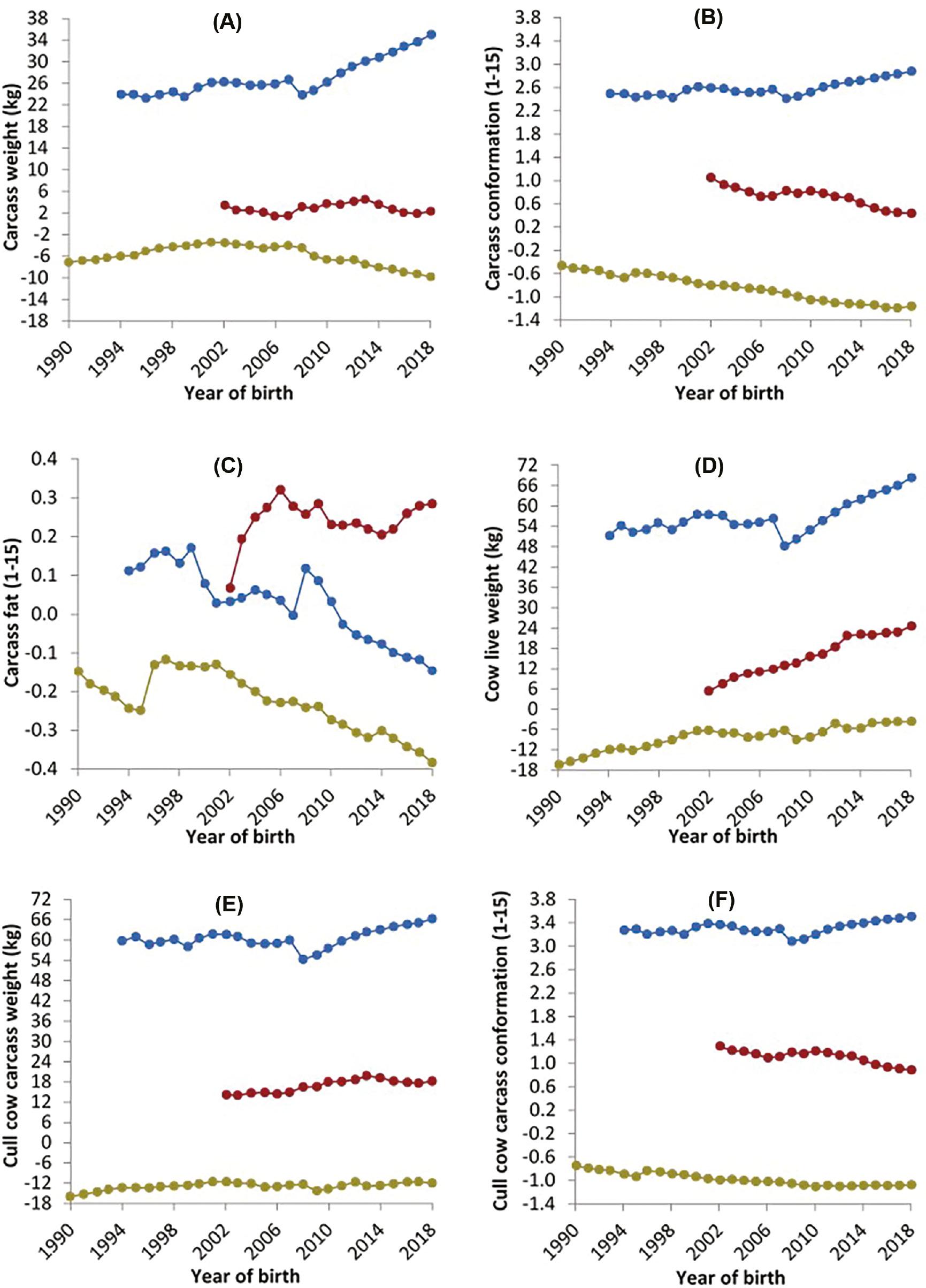
Estimated breeding values for animals from a dairy dam and a dairy sire ( ), a dairy dam and a beef sire (
), a dairy dam and a beef sire ( ) and a beef dam and a beef sire (
) and a beef dam and a beef sire ( ) for (A) carcass weight, (B) carcass conformation, (C) carcass fat, (D) cow live weight, (E) cull cow carcass weight and (F) cull cow carcass conformation for each animal’s birth year.
) for (A) carcass weight, (B) carcass conformation, (C) carcass fat, (D) cow live weight, (E) cull cow carcass weight and (F) cull cow carcass conformation for each animal’s birth year.
For conformation, the EBVs for D×D decreased by 0.02 units per year from 2008 to 2018 irrespective of the breed being adjusted for or not. The genetic trend for conformation showed a decrease of 0.05 units per year for D×B animals; the trend changed when breed covariates were included as there was no genetic gain or deterioration observed per year. An increase in conformation EBVs of 0.05 units per year was observed for B×B animals. When breed covariates were included in the model the conformation EBV trend showed an increase of 0.03 units per year. For carcass fat, the trend was −0.01, 0.00 and −0.02 units per year for D×D, D×B and B×B animals, respectively, for the years 2008–2018. When breed covariates were included in the model the trend for fat was −0.01, −0.01 and −0.01 units per year for D×D, D×B and B×B animals, respectively, for the years 2008–2018.
The genetic trend for cow carcass traits is presented in Figure 1. Cow live weight EBVs increased for D×D, D×B and B×B animals by 0.47, 1.18 and 1.99 kg per year, respectively. The cull cow carcass weight showed an increase of 0.17, 0.12 and 1.18 kg per year, respectively, for D×D, D×B and B×B animals. Cull cow carcass conformation EBVs showed no change for the D×D genotype, a decrease of 0.04 units for the D×B genotype and an increase of 0.04 units for the B×B genotype.
Phenotypic trends
Carcass weight
The phenotypic and genetic trends for steers, young bulls and heifers for carcass weight are presented in Figure 2. For D×D young bulls, the phenotypic trend for carcass weight showed an increase of 1.26 kg per year for the years 2007–2017, while a smaller increase of 0.12 kg was observed for D×D steers. However, in these same animals’ EBVs for the years 2007–2017, a declining trend of 0.50 kg for young bulls and 0.52 kg for steers was noted. The phenotypic trend for carcass weight for the D×B genotype showed an increase of 0.95, 1.22 and 1.48 kg per year for steers, young bulls and heifers, respectively. However, the trend changed and a decrease was observed in these same animals’ EBVs. The genetic trends for these steers, young bulls and heifers showed a decrease of 0.01, 0.01 and 0.02 kg per year, respectively, for the years 2007–2017. A genetic gain of 0.95, 1.19 and 1.10 kg was observed for carcass weight across steers, young bulls and heifers, respectively, for the B×B genotype for the years 2007–2017 inclusive. These animals’ phenotypic carcass weight increased by 1.90 and 2.61 kg for steers and young bulls, respectively, with heifers seeing the largest gain of 3.42 kg per year over the same period.
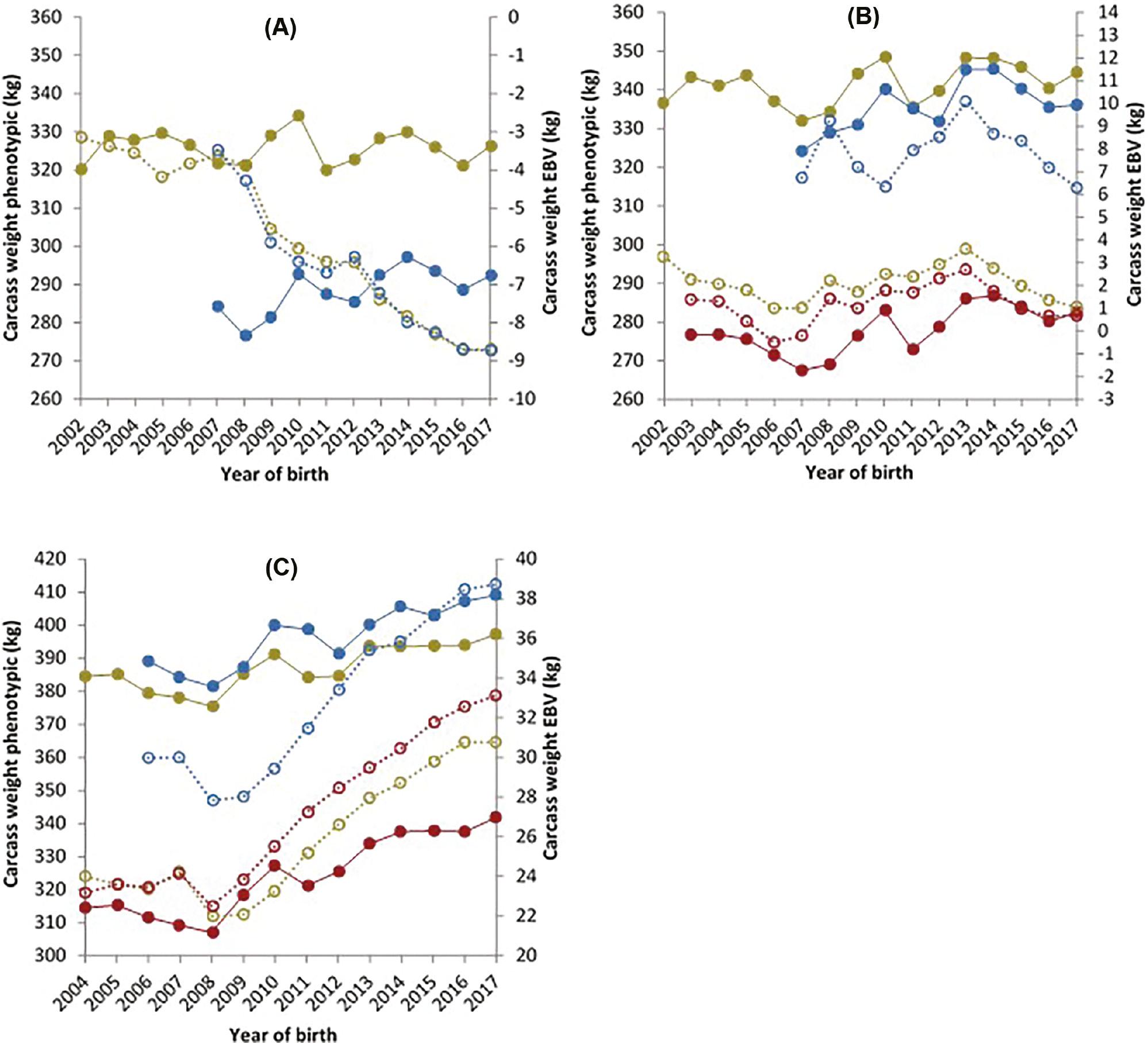
Carcass weight phenotypic data (primary axis) of steers (), heifers () and young bulls () and estimated breeding value’s (secondary axis) of steers ( ), heifers (
), heifers ( ) and young bulls (
) and young bulls ( ) for carcass weight, for animals from (A) a dairy dam and a dairy sire, (B) a dairy dam and a beef sire and (C) a beef dam and a beef sire.
) for carcass weight, for animals from (A) a dairy dam and a dairy sire, (B) a dairy dam and a beef sire and (C) a beef dam and a beef sire.
Carcass conformation
The phenotypic and genetic trends for steers, young bulls and heifers for carcass conformation are presented in Figure 3. Carcass conformation declined both phenotypically and genetically for D×D steers and young bulls. Phenotypically, steers decreased by 0.05 units and young bulls by 0.03 units per year. Their EBVs’ also declined by 0.03 units per year from 2007 to 2017. A similar trend was observed for prime D×B animals. Phenotypic trends for steers, young bulls and heifers all showed a decline of 0.05, 0.05 and 0.03 units per year, respectively. Similarly, the EBVs of these animals also decreased by 0.03, 0.04 and 0.03 units per year for steers, young bulls and heifers, respectively. For B×B animals, gain was observed both genetically and phenotypically for steers, heifers and young bulls. The genetic trend showed an increase of 0.04, 0.05 and 0.05 for steers, young bulls and heifers, respectively, per year from 2007 to 2017. The phenotypic trend showed an increase of 0.03, 0.05 and 0.07 for steers, young bulls and heifers, respectively, for B×B animals.
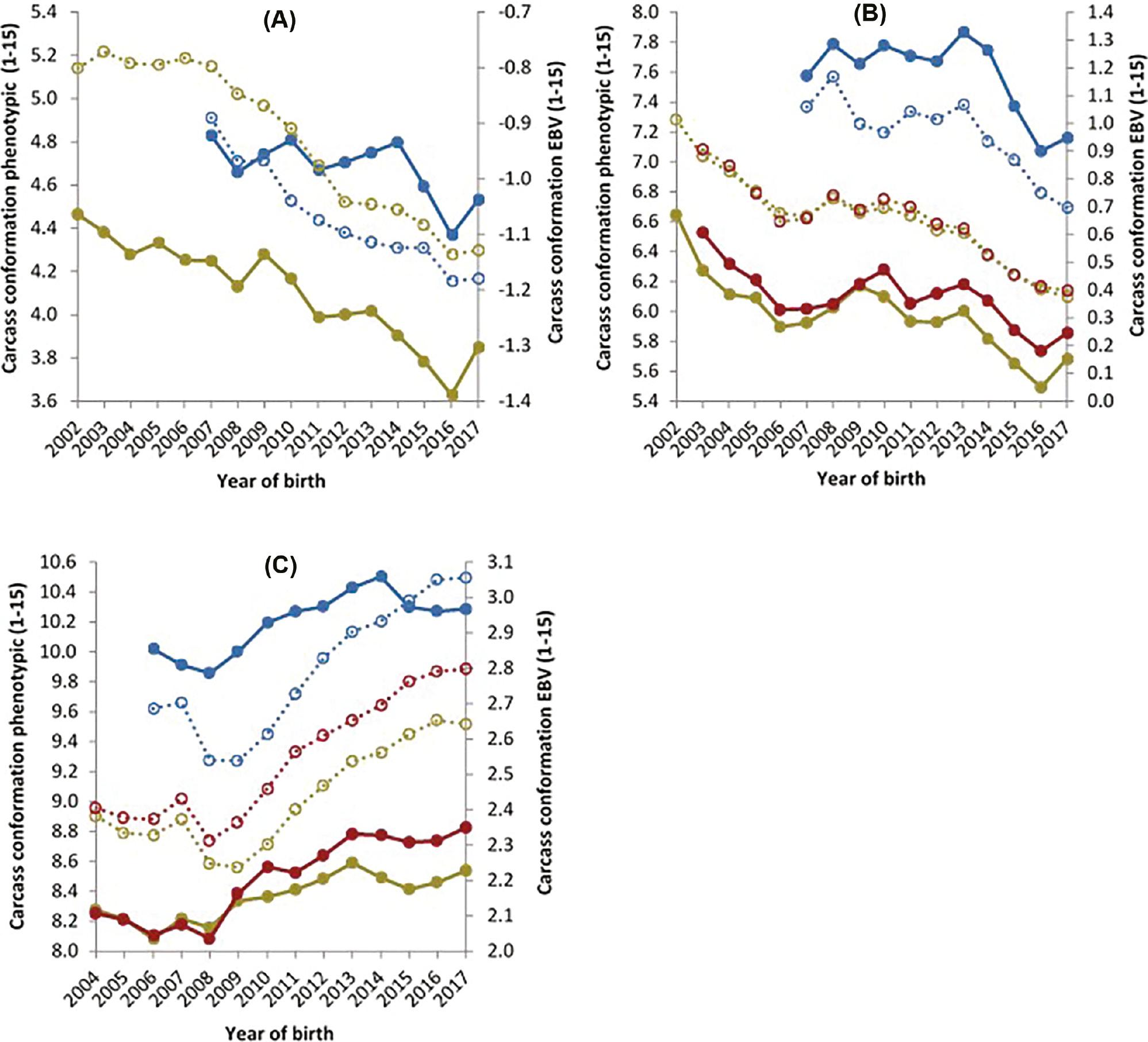
Carcass conformation phenotypic data (primary axis) of steers (), heifers () and young bulls () and estimated breeding value’s (secondary axis) of steers (), heifers () and young bulls () for carcass conformation, for animals from (A) a dairy dam and a dairy sire, (B) a dairy dam and a beef sire and (C) a beef dam and a beef sire.
Carcass fat
The phenotypic and genetic trends for steers, young bulls and heifers for carcass fat are presented in Figure 4. Carcass fat decreased genetically for D×D steers and young bulls (0.02 and 0.01); however, both increased phenotypically (0.02 and 0.05 units) per year, respectively. For D×B heifers, no genetic change for carcass fat was observed, and a decrease in carcass fat EBVs of 0.01 units was seen in both steers and young bulls. Phenotypically, trends for D×B animals showed an increase of 0.03, 0.03 and 0.07 units per year for steers, young bulls and heifers, respectively. Finally, a decrease in carcass fat of 0.03 units per year was observed for B×B animals within all three genders. An increase was seen in the phenotypic data of heifers (0.03 units) and young bulls (0.01 units) for B×B animals and a decrease was observed in steers (0.01 units) per year for the years 2007–2017.
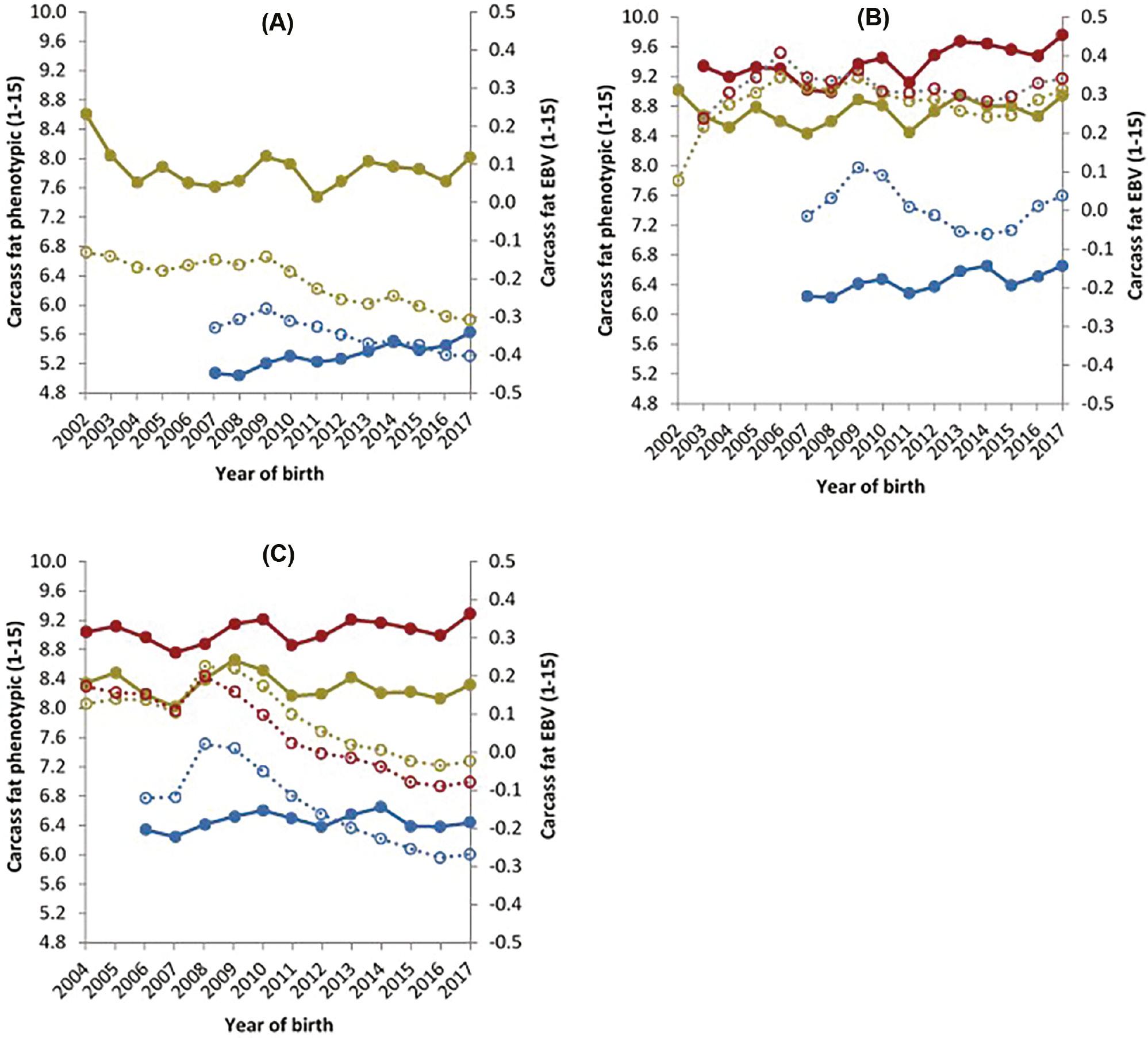
Carcass fat phenotypic data (primary axis) of steers (), heifers () and young bulls () and estimated breeding value’s (secondary axis) of steers (), heifers () and young bulls () for carcass fat, for animals from (A) a dairy dam and a dairy sire, (B) a dairy dam and a beef sire and (C) a beef dam and a beef sire.
Age at slaughter
The phenotypic trend for steers, young bulls and heifers for age at slaughter is presented in Figure 5. There was a consistent reduction in age at slaughter across all steer genotypes. The D×D steers decreased by 2.74 d per year, B×B steers decreased by 3.07 d and D×B steers decreased by 5.07 d per year between the years 2007 and 2017. B×B young bulls exhibited the largest decrease in age at slaughter, with a reduction of 5.32 d per year for the years 2007–2017. A smaller decrease of 0.73 and 1.79 d per year was observed in the D×D and D×B young bulls, respectively. Finally, a decline of 1.72 d and 3.79 d per year was seen in the D×B and B×B heifers, respectively, for the years 2007–2017.

Cow traits
The EBVs (cull cow carcass weight and cull cow carcass conformation) and phenotypic trends (carcass weight and carcass conformation) for cows are presented in Figure 6. D×D cows showed a reduction in phenotypic carcass weight of 0.92 kg per year; however, these same D×D cows’ EBVs showed an increase of 0.13 kg per year for the years 2005–2015. Carcass weight of the D×B cows increased by 0.52 kg genetically and by 1.3 kg phenotypically per year. During the same years B×B cows increased by 0.29 kg per year phenotypically, and their carcass weight EBV showed a similar increase of 0.22 kg per year (Figure 6). With regards to carcass conformation, a similar trend was observed for D×D cow phenotypic and genetic trends, both decreasing by 0.03 units and 0.01 units per year, respectively. No change was observed in the phenotypic and genetic trends for D×B cows’ carcass conformation. The genetic trend for carcass conformation for B×B cows showed a decrease of 0.01 units, per year from 2005 to 2015, while the phenotypic trend showed an increase of 0.01 units (Figure 6).
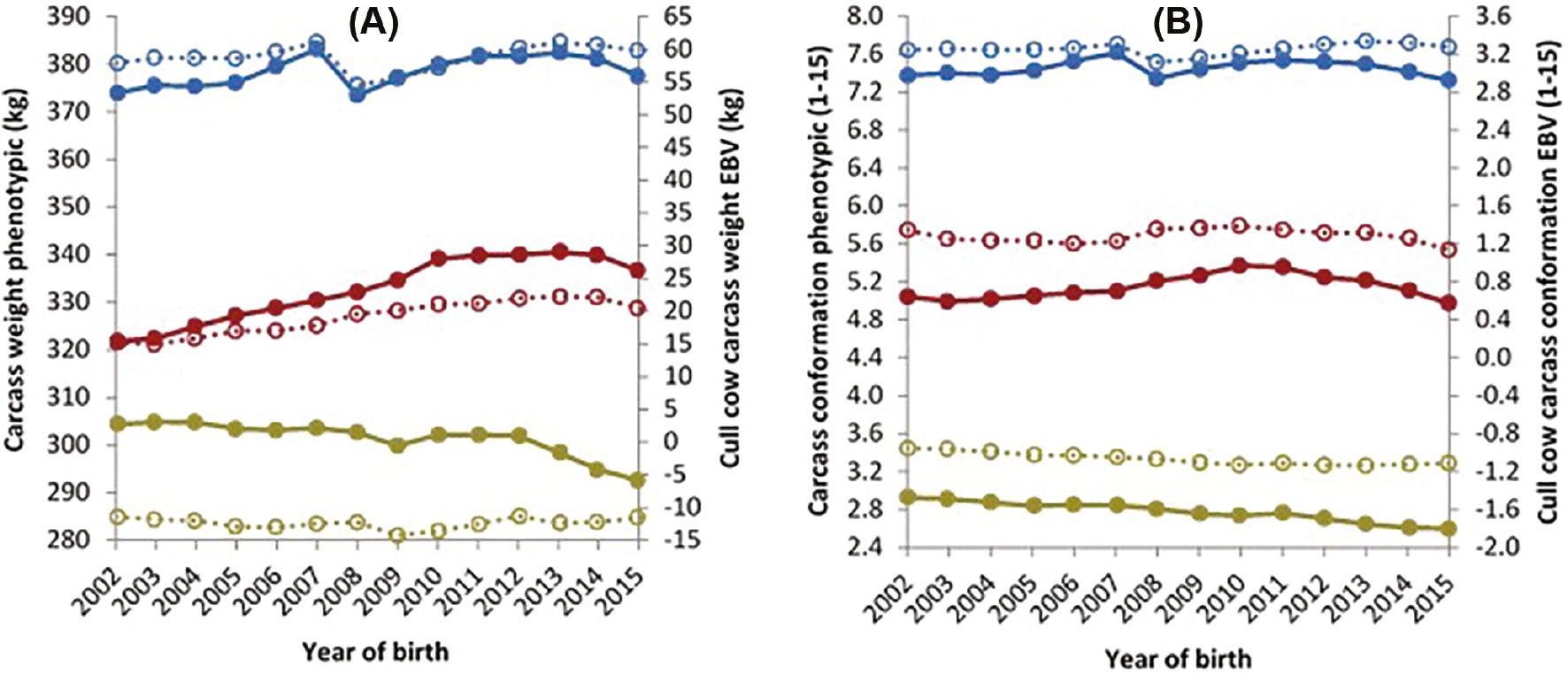
Trends for (A) carcass weight and (B) carcass conformation phenotypic data (primary axis) of cows from a dairy dam and a dairy sire (), a dairy dam and a beef sire () and a beef dam and a beef sire () and (A) cull cow carcass weight and (B) cull cow carcass conformation estimated breeding values (secondary axis) of cows from a dairy dam and a dairy sire (), a dairy dam and a beef sire () and a beef dam and a beef sire ().
Discussion
The purpose of the current study was to assess and quantify the trends in carcass traits in the Irish beef cattle population. The beef industry has set certain standards such as minimum and maximum carcass weights, age restrictions and minimum conformation/fat score requirements (Kenny et al., 2020). One solution to these challenges is to breed for improved carcass merit. The future sustainability of beef production will depend on the genetic ability of animals to meet carcass specifications.
Suckler beef
The favourable improvement in genetic gain for B×B animals’ carcass traits observed in the present study is as expected, as there has always been an emphasis on improving beef traits and these traits have been documented to be highly heritable (Englishby et al., 2016). Prior to the availability of a selection index in 2005, B×B animals were predominately selected based on phenotypic appearance for carcass traits. As observed in the present study, genetic gain for carcass traits was observed for B×B animals from 1990 to 2005. In 2005, ICBF launched three beef selection indices (i.e. Calving Traits Index, Weanling Export Index and the Beef Slaughter Index); however, in 2007, these became sub-indices of the Suckler Beef Index (Evans et al., 2007). The Suckler Beef Index introduced in 2007 for suckler beef farmers included both terminal and maternal traits (Cromie, 2011). Subsequently, the Suckler Welfare Scheme was introduced in 2008 which incentivised farmers to record sires (Cromie et al., 2008). As a result, an increased number of animals had a recorded sire from 2008 to date. For the present study only animals with a recorded sire were retained; thus, the number of B×B animals represented in the genetic trend increased from 2008, with over triple the number of animals in 2008 compared to that in 2007. This shift in the number of animals has caused the fluctuation observed in the trend. In 2013, the Suckler Beef Index was divided into two selection indices. (1) The Terminal Index was designed to breed animals for slaughter, so traits included carcass traits, feed intake and calving traits. (2) The Replacement Index was designed to breed replacement females but also includes carcass traits, as approximately 50% of progeny would be males destined for slaughter. This selection index places a lower relative emphasis (29%) on calf traits (i.e. beef production and calving traits), while it places a higher relative emphasis (71%) on maternal traits (i.e. fertility and milk traits) in comparison to the Terminal Index, which places a full relative emphasis on calf traits. The improvement in all carcass traits for B×B animals accelerated since the introduction of the Terminal and Replacement Indices in 2013, providing the opportunity for beef farmers to select for terminal traits (i.e. carcass weight and conformation) as well as other potentially antagonistically correlated traits such as calving and milk traits. The increase in carcass merit is likely positively influenced by an improvement in data recording at the farm level such as sire recording (Berry et al., 2022). Suckler farmers were incentivised to record data through European Union funded schemes such as the Beef Data Genomics Programme (Vigors, 2017).
Connolly et al. (2016) showed that the Terminal Index improved carcass traits phenotypically even though there is also a large weighting on calving traits within the Terminal Index. Twomey et al. (2020b) found that cows with higher maternal index values had better fertility and weaned heavier calves, while their offspring were able to maintain good carcass performance. This coincides with the genetic improvement seen in B×B animals’ carcass traits as well as similar observations in their phenotypic trends. Nevertheless, the present study does observe an increase in cow live weight in suckler cows which is not a goal in the national maternal selection index, as there is a negative economic weighting on cow live weight in the Replacement Index. However, there is a positive economic weighting on other traits which are correlated to cow live weight (e.g. cull cow carcass weight has a correlation of 0.81 with cow live weight; Berry et al., 2021).
Dairy origin
Unlike B×B animals, D×D animals showed a steady decline in genetic merit for both carcass weight and carcass conformation between the years 2000 and 2018, which is an undesirable trend. The EBI replaced the Relative Breeding Index (RBI) in 2000, which focused solely on genetic advancement in milk production. The RBI was a selection index that consisted of milk yield, fat yield, protein yield and protein percent and it was successful in achieving its goal of producing high-yielding cows. However, the RBI did not include other economically important traits such as fertility and beef (Evans et al., 2002; Berry et al., 2006). In 2005, economic weightings for carcass traits were introduced into the EBI. However, due to the inclusion and selection of other economically important traits such as increased milk production, improved fertility (Berry et al., 2016) and lower cow live weight (Berry et al., 2007), there was a deterioration in the genetic trends for carcass weight and conformation observed in the present study. Carcass traits are not highly ranked in importance to dairy farmers since their main revenue is predominately generated from milk sales. In addition, in most cases, male dairy calves and dairy × beef calves tend to be sold off farm at a young age. Therefore, it is challenging to communicate the significance of the dairy-beef industry to dairy farmers. Nonetheless, dairy and dairy-beef calves must meet a certain carcass standard in order to be profitable for the dairy-beef farmer. Ultimately, it is the dairy farmer who determines the genetics of these animals and will therefore determine how saleable these calves will be in the future. Furthermore, there is a growing concern among consumers about the environmental and ethical impacts of how and where their food is produced (Coleman et al., 2022). It is imperative that the Irish dairy industry ensures all animals have a high quality and purposeful life to ensure Irish dairy is in-line with public values (Ritter et al., 2022). Currently, all trait weightings within the EBI are selected based on a bio-economic model, so economically it makes little sense to increase the weighting for carcass traits as it would reduce the genetic gain of other traits, thereby affecting the profitability of the dairy industry. However, the carcass traits must remain within the EBI to ensure that the inherited beef merit from the dairy cow population does not negatively impact the carcass performance of their progeny destined for beef production. Nevertheless, future changes in markets will determine the value of beef within the bio-economic model and whether or not changes need to be made in the future.
Tools like genomics will allow for faster and more accurate selection in future dairy replacements, making it possible to rapidly and accurately identify elite animals at a younger age. In addition, advances in reproductive technologies, such as sexed semen, along with improved reproductive performance of the dairy herd, will enable dairy farmers to use more beef semen on dairy cows. As a result, a greater proportion of dairy-beef animals will be produced from the dairy herd, thereby reducing the number of surplus calves sired by dairy sires (Murphy et al., 2016). Unfortunately, the genetic trend for D×B animals for carcass weight has shown no improvement. Additional analysis was carried out on carcass weight genetic trends for D×B animals’ sire and dam EBVs. This analysis showed that while the sire carcass weight EBVs of D×B animals increased by 0.14 kg per year, dairy dam carcass weight EBVs of the same animals decreased by 0.4 kg per year, for the years 2008–2018. Thus, the biologically small improvement in the genetic merit for carcass weight of the sire has been offset by a decline in the dairy dam. Similarly, genetic merit for conformation of D×B animals is also declining at the same rate as that of their D×D counterparts. The poorer genetic gain for carcass traits in beef sires used on dairy cows relative to beef cows is likely due to the higher selection emphasis by dairy farmers on calving traits such as short gestation and easier calving, which is known to be negatively correlated with carcass traits (Berry et al., 2020). The availability of a selection index like the DBI, which is an index that ranks beef sires for use on dairy cows to minimise calving difficulty and increase beef merit, was designed to improve the genetic gain for carcass traits of D×B animals (Berry & Ring, 2020).
Breed substitution
While the EBVs utilised in this study come from multi-breed evaluations, it was of interest to see if the changes in genetic trends were due to breed substitution, which may indicate that little or no genetic gain was achieved within individual breeds. For D×D animals, 92% of the breed composition is composed of HO and FR. Over the last 18 years, the proportion of FR has increased from 14% in 2000 to 25% in 2018. However, this breed shift does not represent a breed substitution effect on the genetic trends of carcass weight, carcass conformation and carcass fat. For B×B animals, there has been an increase in the proportion of LM and a small decline in the CH breed composition in recent years (Appendix Figure A1). The genetic gain in carcass weight and conformation of B×B animals was less when breed was accounted for, suggesting that a small proportion of the genetic gain can be attributed to a small breed shift in B×B animals. For D×B animals, the genetic decline in carcass weight and conformation was not observed when adjusted for breed effects; the trend actually reversed for carcass weight. Therefore, the decline in carcass traits of D×B animals is linked to breed introgression rather than an emphasis on poorer genetic merit animals. Dairy farmers are selecting beef bulls with a higher EBV for carcass weight within their chosen breed each year. The breed composition of D×B animals in this study showed an increase in AA and HE usage and decrease in continental breed usage (i.e. BB, CH and LM; Appendix Figure A1). The observed breed substitution for genetic trends in beef sires used on dairy cows is due to the higher selection of breeds that are known for their ease of calving, such as AA and HE. Until recently, there was no selection indices for dairy farmers to select beef sires. The introduction of the DBI which ranks sires across breed will likely help to rectify this decline (Berry et al., 2019).
Reducing age at slaughter
Reducing age at slaughter will be a significant focus in future years to reduce the environmental footprint of beef (Murphy et al., 2017). To achieve this, selection indices will need to ensure animals can reach early slaughter while also meeting required carcass specifications (i.e. weight, fat cover and conformation; Kenny et al., 2020). Even though D×D young bulls and steers were reducing genetically in terms of carcass weight (adjusted to age), phenotypically, these same animals are being slaughtered at a similar carcass weight to previous years but at a younger age than their predecessors. Thus, this suggests that improvements in the management of D×D animals are masking the decline in genetics. The same was the case for D×B animals, albeit not to the same extent. Additionally, the improvement in age is surprising as the present study observed that carcass fat EBV was reducing within each of the three genotypes D×D, D×B and B×B (i.e. animals genetically leaner at a common age), although, phenotypically, each genotype increased. There is a negative economic weighting on fat score in each of the four national selection indices, as animals with lower fat scores have higher meat yields (Berry et al., 2019). It is unknown whether this negative economic weighting has had a negative impact on the length of time required for animals to reach their maturity and, ultimately, their age of slaughter, although the present study suggests that improvements in age at slaughter are still being achieved phenotypically. This improvement in age at slaughter is potentially linked to an intentional production system change on farm, where farmers decided to slaughter animals in a system at a younger age (i.e. shift from a 28-mo slaughter system to a 24-mo system; Ashfield et al., 2013). Berry et al. (2017) proposed breeding for an age at slaughter trait due to the environmental and economic challenges facing the beef industry. Including an age at slaughter trait in national selection indices should contribute to further gains in the phenotypic reduction observed in age at slaughter for the same carcass weight and subcutaneous carcass fatness.
Conclusions and implications
This study gives a thorough insight into the genetic gain of carcass merit using a large dataset of Ireland’s national cattle population. The results indicate that genetic and phenotypic trends for carcass weight, carcass conformation and carcass fat are improving for B×B animals. In contrast, D×D and D×B animals are both declining genetically for carcass weight and carcass conformation. Although the decline was not as apparent phenotypically, effective management and nutrition have masked the genetic decline. Nevertheless, if the genetic trends of D×D and D×B animals continue to decline, it will have a negative impact on the overall beef merit of the national cattle population.

 ), Friesian (
), Friesian ( ), Aberdeen Angus (
), Aberdeen Angus ( ), Belgium Blue (
), Belgium Blue ( ), Charolais (
), Charolais ( ), Hereford (
), Hereford ( ), Limousin (
), Limousin ( ), Simmental (
), Simmental ( ) and for all other breed compositions (
) and for all other breed compositions ( ) for each birth year for animals from (A) dairy dam and dairy sire, (B) dairy dam and beef sire and (C) beef dam and beef sire.
) for each birth year for animals from (A) dairy dam and dairy sire, (B) dairy dam and beef sire and (C) beef dam and beef sire.