
Introduction
Within the United Kingdom (UK) and Ireland, the protein requirement of higher-yielding dairy cows is normally met, in part, through the use of imported protein feeds, including soybean meal and rapeseed meal. However, supply and price volatility, as well as restrictions on the use of genetically modified feeds, have created renewed interest in the use of forage legumes. In addition to their potential to produce high annual yields of dry matter (DM) and crude protein (CP), their ability to fix atmospheric nitrogen (N) reduces the need for N fertiliser, and this can contribute to legume-based systems having a lower carbon footprint (Peyraud et al., 2009).
Red clover (Trifolium pratense L.) is a forage legume of particular interest within the UK and Ireland (Wilkins & Jones, 2000), where it has the potential to produce high annual DM yields (Dale et al., 2014; Clavin et al., 2017). In addition, red clover silage normally has a higher protein content than grass silage (GS) (Dewhurst et al., 2003a). While the protein in red clover may degrade relatively quickly (Dewhurst et al., 2009), the variable presence of polyphenol oxidase (PPO) in red clover can contribute to a lower rate of ruminal protein degradation compared to protein in other forage legumes, such as lucerne (Lee et al., 2004). However, due to its high protein content and the associated reduction in N use efficiency (Dewhurst et al., 2003a; Moorby et al., 2009), red clover silage is often offered in mixtures with GS.
In general, when dairy cows are offered either GS alone or a mixture of red clover silage and GS, the DM intake (DMI) of cows that were offered the mixture increases, with this being often associated with an increase in milk yield, while milk fat and protein content are normally either unaffected or decline (Dewhurst, 2013). However, in the majority of these studies, the GS has been produced from swards that have received moderate or high rates of N fertiliser, and this does not simulate silage produced from mixed swards of grass and red clover, which normally receive either none or only small amounts of N fertiliser. For example, the grass in a grass–red clover sward will have a low N content due to the relatively low amount of N transferred from red clover to the accompanying grass, particularly in the first full harvest year (Dahlin & Stenberg, 2010).
Mixed swards of grass and red clover are often sown on farms in an attempt to increase herbage DM yields, reduce the use of inorganic fertiliser N and increase the nutritive value of the silage produced. In one of the few studies examining their use, Vanhatalo et al. (2006) found that cows that were offered grass–red clover silage had higher milk yields than, but similar contents of DMI, milk fat and milk protein as, cows offered GS. Nevertheless, as in most red clover feeding studies, the silages offered were produced from a single harvest, rather than from successive harvests. In contrast, on commercial dairy farms, silages produced from all harvests within a season are normally offered to livestock, and thus it is important to quantify differences between harvests within a full growing season. This is important as changes in sward structure, maturity and fibre content of the component species in a grass–red clover sward over the course of the growing season will affect the nutritional quality and intake characteristics of the silage and, consequently, milk production and milk composition (Vanhatalo et al., 2009). For example, a regrowth of a grass–red clover sward had a higher CP content and total complement of amino acids than its primary growth (Naadland et al., 2016).
The impact of offering red clover silage to dairy cows does not appear to have been examined previously on the island of Ireland. This is important as research carried out elsewhere may not be directly applicable to Ireland as the maritime climate can lead to unavoidable high humidity or rainfall at harvest, creating sub-optimal conditions for wilting (McEniry et al., 2013), which will affect the silage quality. Consequently, this study was conducted under Irish conditions, and it compared the effects of offering dairy cows grass silage and grass–red clover silages produced from three consecutive harvests over a full production year (harvested in early summer, late summer and autumn) on N and energy utilisation and subsequent animal performance.
Materials and methods
This study was conducted at the Agri-Food and Biosciences Institute (AFBI), Hillsborough, Northern Ireland (54°27′N; 06°04′W). All experimental procedures were conducted under an experimental licence granted by the Department of Health, Social Services & Public Safety for Northern Ireland in accordance with the Animals (Scientific Procedures) Act 1986.
Experimental forages
A perennial ryegrass sward and a red clover–perennial ryegrass mixed sward (approximately 3 ha each) were established during the autumn of 2013. On 3 and 4 September 2013, crops were sown following conventional seedbed preparation using an air seed drill with a mounted spring harrow attachment (Stocks ASg, Cambs, UK; Twose, Ludlow, UK). Seeding rates for red clover (cv. Merviot) and perennial ryegrass (cv. Navan) in the mixed sward were 9.9 kg/ha and 22.2 kg/ha, respectively, while the seeding rate for the perennial ryegrass in the pure sward was 32.1 kg/ha. Neither sward received N fertiliser during establishment. The crops were harvested on three occasions in 2014, namely, 12 June (H1), 7 August (H2) and 2 October (H3). Crops were mown at approximately 14:00 h at each harvest using a mower–conditioner unit (Claas, Bury St Edmunds, UK) and allowed to wilt in the swaths for periods of 48, 72 and 72 h, for H1, H2 and H3, respectively, with the objective of achieving a herbage DM concentration of approximately 270–280 g/kg DM. Swards were placed in windrows using a trailed tractor-driven twin-rotor rower (Claas, Bury St Edmunds, UK) 2–3 h prior to ensilage and subsequently baled using a conventional round baler set at 90% density (Krone, Leeds, UK). The crops were then wrapped using a conventional bale wrapper (McHale, Mayo, Ireland), dispensing plastic film with 17 wraps per bale; each load of bales was weighed on a commercial weighbridge (capacity: 50 t, ± 10 kg) before being stacked. Four samples of each herbage type were collected from the swards immediately in front of the bailer, with these subsequently dried at 85°C for 24 h to determine oven DM (ODM) content. The yields from H1, H2 and H3 for the red clover–GS (RCGS) swards were 4.4, 4.0 and 1.6 t DM/ha, respectively, representing an annual yield of 10.0 t DM/ha. The yields from H1, H2 and H3 for the GS swards were 5.2, 3.4 and 1.8 t DM/ha, respectively, representing an annual yield of 10.4 t DM/ha. Based on visual assessment before mowing, the RCGS swards were estimated to contain approximately 20%, 40% and 60% red clover at each of the harvests H1, H2 and H3, respectively.
In March 2014, both the GS and the RCGS swards received 22 kg N/ha (urea: 46% N). The GS sward received a further 106 kg N/ha (calcium ammonium nitrate: 27% N) in April, followed by 122 kg N/ha after H1 in June and a final application of 65 kg N/ha in August following H2. The RCGS sward received no additional N fertiliser during the growing season.
Animals and housing
Twenty-eight three-way cross (Swedish-Red × Jersey/Holstein-Friesian) dairy cows (22 multiparous, six primiparous), with a mean lactation number of 3.1 and mean days-in-milk of 82 d (s.d., 7.8 d), were used in this experiment. Cows were housed as a single group in a free-stall house with concrete flooring with access to individual cubicles that were fitted with rubber mats, and they were bedded with sawdust twice daily. The cubicle-to-cow ratio was 1:1 at all times, thus meeting the recommendations of the Farm Animal Welfare Committee (FAWC, 1997). The floor area was scraped every 3 h using an automated system.
Treatments
During a 2-wk pre-experimental period, cows were offered GS plus 8.0 kg/d of concentrate via an in-parlour concentrate-feeding system. Cows were allocated to one of two experimental treatments (GS or RCGS) on the final day of the pre-experimental period, with cows within each treatment group balanced for parity (1, 2, 3 and 4) and days in milk; milk yield, milk fat and protein content; live weight (LW); and body condition score (BCS) during the week prior to the start of the study. Throughout the experiment, all cows were offered 8.0 kg/d of a common concentrate through an in-parlour concentrate-feeding system. This was divided into two equal meals, 4.0 kg at each milking. The ingredient composition of the concentrate (in grams per kilogram, fresh basis) was as follows: maize meal, 171; soya bean meal, 132; wheat, 128; rapeseed meal, 125; corn gluten, 124; soya hulls, 101; palm kernel, 99; wheat feed, 45; molaferm (United Molasses, Belfast, UK), 40; lime flour, 9.5; palm oil, 9.5; salt, 8.2; calcined magnesite, 4.1; mineral–vitamin mix, 4; yeast (Actisaf, Lesaffre, Shannon, Ireland), 0.5.
Cows were offered the experimental forages for a 13-wk experimental period. Silages produced from H1, H2 and H3 were offered successively, for periods of 5, 5 and 3 wk, respectively, with the duration of each period broadly reflecting the yields within each harvest of the RCGS sward. Fresh silage was offered daily at approximately 09:00 h (at 107% of the previous day’s fresh weight intake), with uneaten silage removed the following day at approximately 08:00 h. Silage for cows for each treatment was mixed in a feeder wagon (Redrock Vari-Cut-12, Redrock, Armagh, Northern Ireland) for approximately 6 min prior to being deposited in a series of feed boxes mounted on weigh scales. Cows accessed silage in these feed boxes via Calan gates (American Calan Inc., Northwood, NH, USA) linked to an electronic identification system, thus enabling individual cow intakes to be recorded daily. Cows had free access to fresh water at all times.
Cow measurements and sampling of feeds offered
All cows were milked twice daily (between 06:00 and 08:00 h; and between 15:00 and 17:00 h) throughout the experiment using a 50-point rotary milking parlour (Boumatic, Madison, WI, USA), with milk yields automatically recorded at each milking, and the total daily milk yield for each cow for each 24-h period was calculated. Milk samples were taken during two consecutive milkings each week throughout the experiment and analysed for fat, protein and lactose concentrations using an infrared milk analyser (Milkoscan Model 605; Foss Electric, Warrington, UK). A weighted concentration of each constituent was determined for the 24-h sampling period.
LW was recorded twice daily (immediately after each milking) using an automated weighbridge (BioControl, Rakkestad, Norway), and the mean weekly LW for each cow was determined. The BCS of each cow was estimated fortnightly on a 1–5 scale, according to Edmonson et al. (1989), by a trained technician.
Samples of the experimental silages were taken daily throughout the experiment, dried at 85°C for 24 h to determine the ODM content and milled through a mesh sieve with 1-mm apertures. Sub-samples of the dried milled silages were taken twice weekly and bulked for each week, with the bulked samples analysed for neutral detergent fibre (NDF), acid detergent fibre (ADF) and ash concentrations. A fresh sample of each batch of silage was taken weekly and analysed for the following: concentrations of N, ammonia-N, gross energy (GE), fermentation acids (lactic, acetic, propionic, n-butyric and isovaleric acids), ethanol, propanol, metabolisable energy (ME), and pH. The concentrate offered was sampled 3 times weekly, with one sub-sample dried at 85°C for 24 h for determination of ODM content and the dried sample, discarded. A second sub-sample was dried at 60°C for 48 h prior to milling through a 1-mm sieve, bulked for each 14-d period, and analysed for N, NDF, ADF, ash, GE and starch concentrations.
Nutrient utilisation
During the final 8 d of the feeding period within each harvest, four cows from each treatment (the same four cows at each harvest) were used in a nutrient utilisation study. The four cows selected from each treatment group were initially balanced for daily milk yield and LW. All eight cows were transferred into an experimental nutrient utilisation facility (at approximately 09:00 h) and housed in individual stalls, with their lying area comprising a rubber mat. Cows continued to be offered their experimental rations in a feed box located at the front of each stall. The experimental silages were offered ad libitum daily at 09:00 h, while uneaten silage was removed, with refused feed weighed and recorded the following day at 08:00 h. Concentrate (8.0 kg/d) was offered in two equal meals each day (4.0 kg/meal) at 06:30 h and 16:30 h, during milking. This concentrate was offered in plastic feed buckets, which were placed within the feed boxes, and removed after all the concentrates had been consumed at each feeding time. Cows had access to fresh water at all times via a drinker located within each stall. Cows were weighed prior to and on completion of each balance period, with the average LW used in energy utilisation calculations.
Measurements of nutrient utilisation commenced 24 h after cows were placed in the nutrient utilisation facility and comprised a 6-d feeding period (commencing 2 d before the first collection of faeces and urine) and a 6-d total faeces and urine collection period. Faeces were collected in a plastic collection tray (96 cm × 108 cm × 36 cm) placed behind each cow. Urine was collected in a 25-L plastic container via a flexible plastic tube, which was attached to a urine separation system. This was held in position over the vulva by attaching it using Velcro fasteners to a “patch” (Bostik, France) glued on either side of the cow’s tail head. Approximately 300 mL of 50% sulphuric acid was added to each urine collection container when empty to minimise N losses as ammonia. Containers were examined at approximately 20:00 h and, if >60% full approximately, were replaced with an empty container. The total weight of faeces and urine produced during each 24-h collection period was recorded, and a sample of each (0.05 by weight) was retained for subsequent analysis. Faeces and urine samples were stored in a fridge (4°C–6°C) until the final day of the collection period, when the six daily faeces samples and six daily urine samples from each cow were bulked. The single bulked fresh urine sample for each cow was analysed for GE and N concentrations, while the single bulked fresh faeces sample for each cow was analysed for N concentration. A single bulked dried faeces sample for each cow was dried at 85°C for 100 h to determine the ODM, with the dried sample subsequently milled and analysed for ADF, NDF, ash and GE concentrations. A milk sample was taken at each milking, bulked in proportion to yield, with samples bulked for the entire 6-d period and subsequently analysed for GE and N concentrations. Fresh silages offered were analysed daily for ODM, GE and N concentrations, while daily dried samples over the 6-d nutrient utilisation period were bulked and the single bulked sample for each period analysed for ADF, NDF and ash concentrations. The concentrate offered was sampled daily, bulked for the 6-d nutrient utilisation period and its ODM concentration determined. The dry samples were subsequently analysed for GE, NDF, ADF, N and ash concentrations.
In vitro protein degradation kinetics using the DaisyII incubator
Samples of the dry milled silages collected twice weekly were bulked for each harvest and used to determine in vitro ruminal protein degradability using an ANKOM DaisyII incubator (Ankom Technology®, Macedon, NY, USA). Samples (0.5 g) were weighed into F57 filter bags (in triplicate) (which had been pre-rinsed in acetone and allowed to dry) and then heat sealed. The four Daisy jars (2-L capacity) were filled with a warmed (39°C) mixture of solutions A (266 mL) and B (1,330 mL) (1:5 v/v; pH 6.8). The reagents used were as follows: solution A (in 1 L of deionised water), KH2PO4 (10.0 g), MgSO4·7H2O (0.5 g), NaCl (0.5 g), CaCl2·2H2O (0.1 g) and urea (0.5 g); solution B (in 100 mL of deionised water), Na2CO3 (15.0 g) and Nsga2S·9H2O (1.0 g). Rumen fluid was then used to inoculate each Daisy jar. The rumen fluid used was collected 3 h after morning feeding from two Holstein cows fitted with a ruminal cannula, thoroughly mixed together and poured into an insulated bottle prior to transport to the laboratory, where it was then filtered through four layers of cheese cloth. The rumen fluid was then poured into the Daisy jars (400 mL/jar), mixed with solution A and B and the inoculum was again warmed to 39°C. The jars were continuously purged with a stream of CO2 gas to maintain anaerobic conditions.
The in vitro ruminal protein degradation kinetics were studied for each silage type and harvest according to the “gradual addition/all out” schedule (Theodoridou & Yu, 2013), with samples incubated in the jars for 2, 4, 8, 12, 24 and 48 h. The bags assigned to the 48-h incubation period were inserted first, followed 24 h later by those assigned to the 24-h period, with this step repeated for the remaining bags at the appropriate time intervals. Each treatment was contained within separate jars and not mixed with other treatments. The whole process comprised three experimental runs. The number of bags for each treatment and in each experimental run were 2, 2, 2, 2, 6, 4 and 5 for incubation times 0, 2, 4, 8, 16, 24 and 48 h, respectively. The maximum number of bags in each jar at any one time point was 23.
The jars were rotated continuously in the incubation chamber of the DaisyII incubator, and mounted on slow-turning rollers inside the fermentation cabinet, which results in vessel rotation and filtering. After the incubation, the bags were removed from the incubator and rinsed under a cold stream of tap water without detergent to remove excess rumen contents and subsequently dried at 55°C for 48 h and re-weighed. The soluble N fraction was determined by soaking the bags containing the ground samples in warm water (40°C) for 1.5 h and was considered as the proportion of sample N that had disappeared from the bags at T0. The dried samples were kept in a refrigerated room (4°C). Within each treatment, samples from each harvest were subsequently bulked and milled through a 1-mm sieve and analysed for N using the Dumas method (Elementar, Vario Max CN).
Rumen degradation kinetics
In vitro rumen degradation kinetics of CP were determined using the first-order kinetics equation described by Ørskov and McDonald (1979) and modified by Robinson et al. (1986) and Dhanoa (1988) to include lag time:
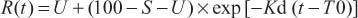
where R(t) is the residue present at t h incubation (%); S is the soluble fraction (%); U is the undegradable fraction (%); D is the potentially degradable fraction (%); T0 is the lag time (h); and Kd is the degradation rate (%/h).
The results were calculated using the NLIN (non-linear) procedure of SAS (SAS Institute, Cary, NC, USA), with iterative least-squares regression (Gauss–Newton method). Based on the non-linear parameters estimated by Equation 1 (S, U and Kd), rumen-degraded feed CP (RDP) and rumen undegraded CP (RUCP) were predicted according to the NRC 2001 model as follows:
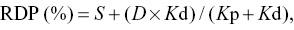
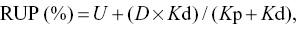
where D = 100 − S − U (%) and Kp is the estimated rate of outflow of digesta from the rumen (%), which was assumed to be 0.06/h, according to Tamminga et al. (1994).
Estimation of the intestinal digestibility of RUCP
Intestinal digestibility of feed RUCP was determined according to the protocol for ruminants (Calamiglia & Stern, 1995). Dried ground rumen residues, containing 15 mg of N, after 16 h of ruminal incubation, were exposed for 1 h to 10 mL of 0.05 mol/L HCl solution containing 1 g/L pepsin. The pH was then neutralised with 0.5 mL of 0.5 mol/L NaOH and 13.5 mL of pH 7.8 phosphate buffer containing 37.5 mg of pancreatin, which were subsequently incubated at 38°C for 24 h. After 24 h incubation, 3 mL of a 100% (wt/vol) trichloroacetic acid (TCA) solution was added to precipitate the undigested proteins. The samples were centrifuged, and the supernatant was analysed for N by the Kjeldahl method (Tecator Kjeltec Auto 2400/2460 Analyzer/Sampler System, Foss). Intestinal digestion of protein was calculated as TCA-soluble N divided by the amount of N in the 16-h residue sample.
Chemical analysis of feed, faeces and urine samples obtained during the study
Nitrogen concentrations of fresh samples were determined using the Kjeldahl method, while N concentrations of dried samples were determined using the Dumas method (Elementar, Vario Max CN). Concentrations of NDF and ADF were determined using a Fibertec analyser (Fibertec FT122; Foss, Hillerød, Denmark) based on the method of Van Soest (1976). Ash concentrations were determined following combustion in a muffle furnace at 550°C for approximately 10 h. Starch concentrations were determined using a Megazyme Kit (Megazyme International, Bray, Ireland). GE concentrations of fresh silage, dry concentrate and faeces, and of freeze-dried samples of milk and urine, were determined using a bomb calorimeter (Parr 6400 Bomb Calorimeter; Parr Instrument Co., Moline, IL, USA). Silage fermentation acids, ethanol and propanol were determined using single-column gas–liquid chromatography (Varian Star 3400 CX GC, equipped with a flame ionisation detector), where samples were injected on column. Ammonia nitrogen concentrations in silage and the silage pH were determined as described by Steen (1989). The ME concentration of the fresh silages was determined using near-infrared reflectance spectroscopy (NIRS), using a calibration developed for GSs, according to Park et al. (1998).
Statistical analysis
Differences in the mean chemical composition of the GS and RCGS within each of the harvests H1–H3 were tested for significance by Student’s two-tailed t-test using Microsoft Excel (2013). In vitro protein degradation mean values were tested for significance by two-way analysis of variance (ANOVA), with “runs” considered as replicates. Mean data for DMI, milk yield, milk composition, LW and BCS, as recorded during the period when each batch of silage was offered and over the entire experimental period, were analysed using ANOVA. When significant, appropriate pre-experimental variables were included as covariates in the model when analysing the corresponding dependant variables (pre-experimental milk yield for milk yield, pre-experimental milk composition values for milk fat, protein, fat-plus-protein and mean daily milk yields; pre-experimental LW for mean LW; pre-experimental BCS for mean and final BCS; pre-experimental LW for forage, concentrate and total DMI). The effects of forage type on the total diet digestibility coefficients, as well as on the N and GE parameters, were examined using a two-way ANOVA. Mean values for protein degradation were analysed using one-way ANOVA. All data were analysed using GenStat (16th edition; VSN International Ltd, Hemel Hempstead, UK).
Results
Chemical composition and in vitro protein degradability of silages
The concentrate offered had the following values: DM, 894 (s.d., 3.1) g/kg; CP, 217 (s.d., 2.54) g/kg DM; ADF, 159 (s.d., 6.7) g/kg DM; NDF, 325 (s.d., 4.0) g/kg DM; ash, 76 (s.d., 1.5) g/kg DM; GE, 18.0 (s.d., 0.04) MJ/kg DM; and starch, 229 (s.d., 8.7) g/kg DM. The volatile-corrected ODM content of RCGS was higher than that of GS at H1 (P = 0.014), while the reverse was true at H3 (P = 0.005) (Table 1). At H1, forage CP (Table 1) tended to be higher in GS than in RCGS (P = 0.074), while at H3, it was lower in GS than in RCGS (P = 0.006). At H1, both the ADF and NDF of GS were higher than in RCGS (P = 0.001, P = 0.002, respectively). At H2, the GS had a higher NDF content than RCGS (P = 0.002), while the reverse tended to be the case for ADF content (P = 0.059). Contents of acetic acid, propionic acid and n-butyric acid were higher in GS than in RCGS at H1 (P = 0.005, 0.017 and 0.001, respectively) and lower in GS than in RCGS at H3 (P < 0.001, <0.005 and <0.043, respectively). Acetic acid content was higher in RCGS than in GS at H2 (P = 0.034). The RCGS samples tended to have a lower lactic acid content than GS samples at H1 (P = 0.087), while at H3, it was higher in RCGS than in GS (P = 0.044). Ammonia nitrogen was higher, respectively, in GS than in RCGS at H1 (P = 0.002), and pH was lower in GS than in RCGS at H2 (P = 0.014). The ME contents of the two silages were not significantly different at either H1 or H2, while tending (P = 0.088) to be higher for the GS at H3.
Chemical composition of grass silage (GS) and red clover–grass silage (RCGS) offered during the 13-wk experimental period at primary growth (H1) or the first (H2) or second (H3) regrowth
| H1 | H2 | H3 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| ||||||||||||
| GS | RCGS | s.e. | P-value | GS | RCGS | s.e. | P-value | GS | RCGS | s.e. | P-value | |
| VCODM (g/kg) | 218 | 299 | 18.3 | 0.014 | 188 | 204 | 5.8 | 283 | 216 | 16.0 | 0.005 | |
| Crude protein (g/kg DM) | 132 | 98 | 11.5 | 137 | 148 | 6.03 | 158 | 216 | 13.8 | 0.006 | ||
| Ash (g/kg DM) | 82 | 75 | 2.7 | 90 | 109 | 2.9 | 0.001 | 87 | 105 | 5.2 | 0.002 | |
| ADF (g/kg DM) | 371 | 344 | 3.8 | <0.001 | 362 | 383 | 5.6 | 0.023 | 262 | 269 | 6.1 | |
| NDF (g/kg DM) | 631 | 596 | 5.7 | 0.002 | 625 | 565 | 13.2 | 0.010 | 482 | 462 | 9.9 | |
| pH | 4.3 | 4.2 | 0.09 | 4.4 | 4.7 | 0.07 | 0.014 | 4.5 | 4.7 | 0.06 | ||
| Lactic acid (g/kg DM) | 97.1 | 67.3 | 14.13 | 83.0 | 31.1 | 13.5 | 0.029 | 80.5 | 96.9 | 11.50 | ||
| Acetic acid (g/kg DM) | 22.6 | 12.5 | 1.85 | 0.005 | 24.2 | 38.7 | 1.99 | 0.001 | 25.9 | 36.3 | 3.36 | <0.001 |
| Propionic acid (g/kg DM) | 2.4 | 0.7 | 0.39 | 0.017 | 1.9 | 3.8 | 0.51 | 0.036 | 0.5 | 3.7 | 0.78 | 0.005 |
| n-Butyric acid (g/kg DM) | 14.1 | 3.6 | 1.33 | 0.001 | 15.3 | 21.6 | 3.24 | 0.0 | 3.6 | 0.98 | 0.043 | |
| Propanol (g/kg DM) | 0.4 | 0.0 | 0.20 | 0.0 | 0.0 | 0.00 | 0.0 | 0.1 | 0.07 | |||
| Ethanol (g/kg DM) | 1.3 | 0.3 | 2.04 | 9.9 | 2.1 | 1.21 | 0.006 | 7.7 | 6.6 | 2.51 | ||
| Ammonia (g/kg total N) | 17.4 | 9.2 | 1.25 | 0.002 | 18.7 | 17.0 | 0.89 | 10.3 | 14.3 | 1.04 | 0.027 | |
| Metabolisable energy (MJ/kg DM) | 10.4 | 10.5 | 0.14 | 10.2 | 9.8 | 0.20 | 10.5 | 9.8 | 0.19 | |||
VCODM = volatile-corrected oven dry matter; ADF = acid detergent fibre; NDF = neutral detergent fibre.
Across the six forages offered, there were no differences in D (potentially degradable fraction) or K d (degradation rate) (Table 2). The potentially soluble fraction (S) was significantly higher in GS than in RCGS at H3 only. The undegradable fraction (U) was significantly higher in RCGS than in GS at each of H2 and H3. The RCGS had a significantly higher RUCP at H2 and H3, while the reverse was true for effectively degraded CP (EDCP). While in vitro intestinal digestibility of CP (IVCPD) was higher with RCGS at H1, it was unaffected by silage type at either H2 or H3. The in vitro intestinal digestibility of the undegradable CP in the rumen (IVCPDCP) was higher with RCGS at H3 but did not differ at either H1 or H2 (Table 2).
In vitro ruminal protein degradation kinetics of grass silage (GS) or red clover–grass silage (RCGS) produced from the primary growth (H1), the first regrowth (H2), or the second regrowth (H3)
| GS H1 | RCGS H1 | GS H2 | RCGS H2 | GS H3 | RCGS H3 | s.e. | P-value | |
|---|---|---|---|---|---|---|---|---|
| S (%CP) | 53.4a | 55.9a | 34.3b | 27.3b | 63.6a | 32.0b | 4.76 | 0.017 |
| D (%CP) | 26.4 | 27.9 | 35.1 | 35.4 | 28.6 | 34.2 | 3.99 | |
| U (%CP) | 20.2c | 16.1c | 30.6b | 37.3a | 7.8d | 33.8a,b | 1.64 | <0.001 |
| Kd (% h) | 15.0 | 11.2 | 20.6 | 9.1 | 8.2 | 10.9 | 6.20 | |
| RUCP (%CP) | 29.3c | 25.8c,d | 41.7b | 51.4a | 19.8d | 46.4a,b | 2.30 | 0.001 |
| EDCP (%CP) | 70.7b | 74.2a,b | 58.3c | 48.6d | 80.2a | 53.6c,d | 2.30 | 0.001 |
| IVCPD (%) | 71.1b | 84.2a | 65.7c | 62.9c,d | 55.6e | 58.9d,e | 1.35 | <0.001 |
| IVCPDCP (%) | 20.9b | 21.7b | 27.5a,b | 32.4a | 11.0c | 27.6a,b | 2.14 | 0.009 |
Mean values with different letters a–e within the same row differ significantly.
CP = crude protein; S = potential soluble fraction in the in vitro ruminal incubation; D = potentially degradable fraction in the in vitro ruminal incubation; U = undegradable fraction; K d = degradation rate; RUCP = rumen undegraded CP; EDCP = effectively degraded CP; IVCPD = in vitro intestinal digestibility of CP; IVCPDCP = in vitro intestinal digestibility of the undegradable CP in the rumen.
Cow performance
Silage DMI was lower in GS than in RCGS treatments in H1 (P < 0.001) and H2 (P = 0.006) and across the entire experimental period (H1–H3: P < 0.001; Table 3). Differences in total DMI were similar. Milk yield was unaffected by silage type at H1 (P > 0.05), tended to be lower for cows offered GS than RCGS at H2 (P = 0.093), while being significantly lower for those offered GS at H3 (P = 0.002). Milk fat concentration was unaffected by treatment at H1 and H2, but it was higher in GS than in RGCS at H3 (P < 0.001). Similarly, milk protein concentration was unaffected by treatment at H1, tended to be significantly lower for cows offered RCGS than GS at H2 (P = 0.080) and was lower for cows offered RCGS than GS at H3 (P < 0.001). Milk fat + protein yield was unaffected by treatment at either H1 or H2, but it was lower with cows offered RCGS than GS at H3 (P = 0.001). Cows offered RCGS were heavier (P = 0.049) and had a higher BCS (P = 0.042) than those offered GS at H1, while neither LW nor BCS was affected by treatment at either H2 or H3.
Performance of dairy cows offered either grass silage (GS) or red clover–grass silage (RCGS) from primary growth (H1), first regrowth (H2) or second regrowth (H3)
| Treatment | ||||
|---|---|---|---|---|
|
| ||||
| GS | RCGS | s.e. | P-value | |
| H1 | ||||
| Silage DMI (kg/d) | 8.8 | 11.7 | 0.28 | <0.001 |
| Total DMI (kg/d) | 15.9 | 18.8 | 0.28 | <0.001 |
| Milk yield (kg/d) | 24.9 | 25.5 | 0.36 | |
| Milk fat (g/kg) | 47.3 | 47.3 | 1.10 | |
| Milk protein (g/kg) | 31.6 | 32.3 | 0.41 | |
| Milk lactose (g/kg) | 46.5 | 46.9 | 0.33 | |
| Fat + protein yield (kg/d) | 2.69 | 2.72 | 0.094 | |
| Average live weight (kg) | 549 | 558 | 2.9 | 0.049 |
| Average body condition score | 2.56 | 2.68 | 0.037 | 0.042 |
| H2 | ||||
| Silage DMI (kg/d) | 9.2 | 10.4 | 0.28 | 0.006 |
| Total DMI (kg/d) | 16.4 | 17.5 | 0.28 | 0.006 |
| Milk yield (kg/d) | 22.1 | 23.1 | 0.44 | |
| Milk fat (g/kg) | 45.3 | 45.2 | 0.98 | |
| Milk protein (g/kg) | 32.0 | 30.8 | 0.47 | |
| Milk lactose (g/kg) | 45.4 | 45.4 | 0.47 | |
| Fat + protein yield (kg/d) | 2.47 | 2.43 | 0.090 | |
| Average live weight (kg) | 547 | 550 | 4.3 | |
| Average body condition score | 2.52 | 2.62 | 0.051 | |
| H3 | ||||
| Silage DMI (kg/d) | 12.3 | 11.5 | 0.45 | |
| Total DMI (kg/d) | 19.5 | 18.7 | 0.45 | |
| Milk yield (kg/d) | 21.8 | 24.7 | 0.58 | 0.002 |
| Milk fat (g/kg) | 47.8 | 43.1 | 0.82 | <0.001 |
| Milk protein (g/kg) | 35.1 | 31.4 | 0.45 | <0.001 |
| Milk lactose (g/kg) | 44.5 | 45.2 | 0.55 | |
| Fat + protein yield (kg/d) | 2.74 | 2.41 | 0.085 | |
| Average live weight (kg) | 569 | 559 | 5.8 | |
| Average body condition score | 2.53 | 2.58 | 0.051 | |
| H1–3 | ||||
| Silage DMI (kg/d) | 9.5 | 11.1 | 0.26 | <0.001 |
| Total DMI (kg/d) | 16.7 | 18.3 | 0.26 | <0.001 |
| Milk yield (kg/d) | 23.4 | 24.4 | 0.39 | |
| Milk fat (g/kg) | 46.8 | 45.8 | 0.82 | |
| Milk protein (g/kg) | 32.3 | 31.5 | 0.41 | |
| Milk lactose (g/kg) | 45.7 | 46.0 | 0.40 | |
| Fat + protein yield (kg/d) | 2.62 | 2.56 | 0.080 | |
| Average live weight (kg) | 552 | 555 | 3.71 | |
| Average body condition score | 2.54 | 2.64 | 4.21 | |
| End of study live weight (kg) | 569 | 559 | 5.8 | |
| End of study body condition score | 2.52 | 2.56 | 0.050 | |
DMI = DM intake.
Nutrient utilisation
None of the digestibility parameters examined differed between GS and RCGS at H1 (P > 0.05) (Table 4). Dry matter digestibility, organic matter digestibility and digestible organic matter in DM were higher for GS than for RCGS at H2 (P < 0.001, P = 0.003 and P < 0.001, respectively) and H3 (P = 0.006, 0.007 and 0.002, respectively). Digestibility of ADF and NDF was higher for GS than for RCGS at H2 (P < 0.001 and P = 0.017, respectively) and H3 (P = 0.009 and 0.037, respectively).
Digestibility of components of DM consumed by dairy cows offered either grass silage (GS) or red clover–grass silage (RCGS) produced from primary growth (H1), first regrowth (H2) or second regrowth (H3)
| Treatment | ||||
|---|---|---|---|---|
|
| ||||
| GS | RCGS | s.e. | P-value | |
| H1 | ||||
| Digestibility (kg/kg) | ||||
| DM | 0.74 | 0.75 | 0.006 | |
| Organic matter | 0.75 | 0.76 | 0.006 | |
| Digestible organic matter in DM | 0.69 | 0.70 | 0.005 | |
| ADF | 0.71 | 0.71 | 0.006 | |
| NDF | 0.69 | 0.70 | 0.007 | |
| H2 | ||||
| Digestibility (kg/kg) | ||||
| DM | 0.74 | 0.68 | 0.006 | 0.001 |
| Organic matter | 0.75 | 0.70 | 0.008 | 0.003 |
| Digestible organic matter in DM | 0.69 | 0.63 | 0.007 | 0.001 |
| ADF | 0.75 | 0.63 | 0.010 | 0.001 |
| NDF | 0.69 | 0.55 | 0.031 | 0.017 |
| H3 | ||||
| Digestibility (kg/kg) | ||||
| DM | 0.76 | 0.71 | 0.009 | 0.006 |
| Organic matter | 0.77 | 0.73 | 0.007 | 0.007 |
| Digestible organic matter in DM | 0.70 | 0.65 | 0.007 | 0.002 |
| ADF | 0.78 | 0.72 | 0.010 | 0.009 |
| NDF | 0.71 | 0.66 | 0.012 | 0.037 |
ADF = acid detergent fibre; NDF = neutral detergent fibre.
Faecal N/N intake was higher for RCGS than for GS at H1 (P = 0.019) but did not differ between silages at H2 and H3, while urinary N/N intake was higher for GS than for RCGS at H1 (P = 0.008; Table 5). Manure N/N intake was higher for GS at H1 (P = 0.005), while the reverse was observed at H2 (P = 0.002). Milk N/N intake was higher for GS than for RCGS at H2 and H3 (P = 0.007 and 0.020, respectively). Retained N/N intake was higher for GS than for RCGS at H2 (P = 0.035), but it was unaffected at H1 and H3. At H1, faecal N/manure N was higher in RCGS than in GS (P = 0.010), while urinary N/manure N was higher for GS than for RCGS (P = 0.010).
Nitrogen utilisation of dairy cows offered either grass silage (GS) or red clover–grass silage (RCGS) produced from primary growth (H1), first regrowth (H2) or second regrowth (H3)
| Treatment | ||||
|---|---|---|---|---|
|
| ||||
| GS | RCGS | s.e. | P-value | |
| H1 | ||||
| N utilisation (g/g) | ||||
| Faecal N/N intake | 0.31 | 0.37 | 0.014 | 0.019 |
| Urinary N/N intake | 0.33 | 0.22 | 0.019 | 0.008 |
| Manure N/N intake | 0.63 | 0.59 | 0.007 | 0.005 |
| Milk N/N intake | 0.28 | 0.29 | 0.016 | |
| Retained N/N intake | 0.09 | 0.12 | 0.017 | |
| Faecal N/manure N | 0.48 | 0.62 | 0.027 | 0.010 |
| Urinary N/manure N | 0.52 | 0.38 | 0.027 | 0.010 |
| H2 | ||||
| N utilisation (g/g) | ||||
| Faecal N/N intake | 0.29 | 0.32 | 0.010 | |
| Urinary N/N intake | 0.26 | 0.33 | 0.019 | |
| Manure N/N intake | 0.56 | 0.65 | 0.012 | 0.002 |
| Milk N/N intake | 0.26 | 0.23 | 0.005 | 0.007 |
| Retained N/N intake | 0.19 | 0.13 | 0.015 | 0.035 |
| Faecal N/manure N | 0.53 | 0.50 | 0.023 | |
| Urinary N/manure N | 0.47 | 0.50 | 0.023 | |
| H3 | ||||
| N utilisation (g/g) | ||||
| Faecal N/N intake | 0.42 | 0.41 | 0.013 | |
| Urinary N/N intake | 0.44 | 0.47 | 0.021 | |
| Manure N/N intake | 0.86 | 0.87 | 0.023 | |
| Milk N/N intake | 0.28 | 0.24 | 0.009 | 0.020 |
| Retained N/N intake | −0.14 | −0.11 | 0.018 | |
| Faecal N/manure N | 0.50 | 0.46 | 0.021 | |
| Urinary N/manure N | 0.50 | 0.54 | 0.021 | |
Gross energy digestibility and digestible energy (DE)/GE were higher with GS than with RCGS at H2 (P = 0.003 and 0.002, respectively) but not at H1 and H3 (Table 6). Milk energy/ME intake tended to be higher with GS than with RCGS at H1 (P = 0.094) while being higher at H3 (P < 0.001). At H2, the proportion of digestible energy intake (DEI) that was lost as faecal and urinary energy was higher in RCGS than in GS (P = 0.002 and 0.003, respectively), while at H3, faecal energy/DEI was unaffected by treatment, while urine energy/DEI was higher with GS (P = 0.032).
Gross energy digestibility and energy utilisation of dairy cows offered either grass silage (GS) or red clover–grass silage (RCGS) produced from primary growth (H1), first regrowth (H2) or second regrowth (H3)
| Treatment | ||||
|---|---|---|---|---|
|
| ||||
| GS | RCGS | s.e. | P-value | |
| H1 | ||||
| GE digestibility | 0.72 | 0.73 | 0.006 | |
| Energy utilisation (MJ/MJ) | ||||
| DE/GE | 0.73 | 0.73 | 0.006 | |
| Milk energy/ME | 0.40 | 0.34 | 0.021 | |
| Faeces energy/DEI | 0.38 | 0.37 | 0.011 | |
| Urine energy/DEI | 0.03 | 0.03 | 0.003 | |
| H2 | ||||
| GE digestibility | 0.73 | 0.67 | 0.008 | 0.003 |
| Energy utilisation (MJ/MJ) | ||||
| DE/GE | 0.73 | 0.67 | 0.008 | 0.002 |
| Milk energy/ME | 0.39 | 0.40 | 0.009 | |
| Faeces energy /DEI | 0.37 | 0.49 | 0.015 | 0.002 |
| Urine energy /DEI | 0.03 | 0.06 | 0.004 | 0.003 |
| H3 | ||||
| GE digestibility | 0.66 | 0.67 | 0.009 | |
| Energy utilisation (MJ/MJ) | ||||
| DE/GE | 0.66 | 0.67 | 0.009 | |
| Milk energy/ME | 0.51 | 0.42 | 0.011 | 0.001 |
| Faeces energy /DEI | 0.51 | 0.49 | 0.015 | |
| Urine energy /DEI | 0.09 | 0.07 | 0.004 | 0.032 |
DEI = digestible energy intake; GE = gross energy; ME = metabolisable energy.
Discussion
This study examined the performance and nutrient utilisation of dairy cows offered silages produced from three successive harvests of a red clover–perennial ryegrass sward and a perennial ryegrass sward. While previous studies have examined the impact of mixing red clover silages and GS on cow performance post-harvest, few have examined silages produced from mixed swards across multiple harvests, as would be commonly practised under commercial farm conditions.
Sward production
The annual DM production from the RCGS sward (10.0 t DM/ha) in the current study was similar to that recorded in a field-scale study by Castle & Watson (1974) in western Scotland (10.9 t DM/ha). However, these yields were considerably lower than the 14.7 t DM/ha reported by Roberts et al. (1990) from a mixed red clover–grass sward in its first harvest year in southern Scotland.
While pure red clover crops have been reported to fix up to 250 kg N/ha (Smith et al., 1985), less is known about the N-fixing potential of red clover within grass swards. In the current study, the total annual DM produced from the mixed sward (which received only 22 kg N, in early spring) was only 0.4 t DM/ha less than that produced from the pure perennial ryegrass sward (10.4 t DM/ha), which received a total of 315 kg N/ha. This suggests that the amount of N the red clover component contributed to the sward was equivalent to a grass sward receiving approximately 263 kg N/ha, assuming that an additional 0.4 t DM/ha was produced by 30 kg N/ha (Morrison et al., 1980), that is 315 – 30 – 22 kg N/ha.
Silage characteristics
Within each harvest, both the grass and the grass–red clover silages were produced under the same climatic conditions, with both swards wilted for similar periods. However, silage DM contents indicate that wilting conditions were poor within the current study, as occurs frequently in Ireland. In general, red clover tends to have higher moisture content than grass at any given time during the growth cycle (Hynes et al., 2018), and this likely accounts for the lower DM of RCGS than GS at H3, the former estimated to contain approximately 60% red clover. In contrast, the higher DM content of the RCGS than GS at H1 is likely to reflect the lower total yield of crop, resulting in more rapid drying than the heavier GS crop (Wright et al., 1997) and the low red clover proportion with this crop at first harvest (visually estimated to be approximately 20%).
The CP content of a pure red clover sward has been found to increase progressively with harvest. For example, in a trial in Ireland, CP increased from 155 g/kg at the first to 263 g/kg at the fourth harvest (Clavin et al., 2017). The increase in the CP content of the RCGS silages (98, 148 and 216 g/kg DM for H1, H2 and H3, respectively) reflects the increasing CP content of the swards as red clover content increased with progress through the growing season, in addition to the seasonal increase in CP content as found by Clavin et al. (2017). This compares to a CP content of the GS at H3 of 158 g/kg DM, despite the much higher rate of N fertiliser applied to GS than to RCGS.
The presence of vegetative red clover in the sward is likely to have contributed to the lower concentrations of ADF and NDF in RCGS than in GS at H1. The vegetative state of both sward types at H3 would have been responsible for the lower ADF and NDF contents at this harvest compared to H1 and H2. The higher pH for red clover silage than GS at H2 is similar to the 0.7-unit difference between the two silage types observed by Moorby et al. (2009). While this can be attributed to the high buffering capacity of red clover compared to perennial ryegrass (King et al., 2012), the reason that no such effect was observed at H3 is unclear. While red clover silages tend to have higher concentrations of volatile fatty acids (VFAs) than GS (Moorby et al., 2009), no consistent trends in VFA content between RCGS and GS were observed in the current study, possibly due to the lower DM content of RCGS than GS within harvests.
Although levels are variable, red clover PPO can oxidise phenols to produce quinones that bind with specific sites on some proteins, reducing proteolysis in the rumen (Theodorou et al., 2006). The presence of PPO may explain the higher proportion of CP that was undegraded in the rumen in RCGS compared to GS at H2 and H3 when assessed by the in vitro method. The digestibility of undegradable protein declined with successive harvests, thus suggesting less N to be readily available for intestinal absorption. Johansen et al. (2017) found total tract digestibility of CP in a perennial ryegrass–red clover silage to be slightly but significantly lower than that of the corresponding GS, but the rumen degradation rate of CP in red clover was faster than in perennial ryegrass. In our study, although total degradation of protein in RGCS was lower than in GS, there was no difference in degradation rate between the two silages.
Intake and performance
Over the entire experimental period, forage DMIs with RCGS were 1.6 kg/d higher than for GS. However, this effect was not consistent across harvests, with intakes with RCGS being higher at H1 and H2 but not at H3. In general, intakes increase with red clover inclusion in the diet. For example, Moorby et al. (2009), with a diet containing a mixture of red clover and GS (one third red clover on a DM basis) and Kuoppala et al. (2009), with a diet containing equal mixtures of red clover and GS on a DM basis, found intakes to be 1.1 and 2.0 kg DM/d higher, respectively, than with pure GS. Halmemies-Beauchet-Filleau et al. (2012) also found an increase in forage DMI when red clover silage was mixed with GS, compared to a GS-only diet.
While in vivo digestibility and in vitro degradability characteristics of the two silage types did not differ for any parameter at H1, RCGS had a much higher DM content at H1 than at subsequent harvests, potentially contributing to the higher DMI at this harvest. In addition, RCGS had a lower fibre content than GS, reflecting the visibly less mature state of the crop at harvest than at later harvests. However, the higher intakes with RCGS than with GS at H2 occurred despite the two silages having similar degradability characteristics and DM content and the RCGS having a lower digestibility coefficient for each of the parameters measured. Nevertheless, the NDF content of the RCGS was substantially lower than for the GS. In contrast, at H3, although red clover content was at its highest, DMI of RCGS was similar to that of GS at this harvest. As with H2, digestibility of RCGS was lower than that of GS; however, the similar NDF contents between the two silages may have been a contributing factor (Weisbjerg & Soegaard, 2008), combined with the higher DM content of the GS. A similar effect was observed by Halmemies-Beauchet-Filleau et al. (2014).
Dry matter intake is partially inversely related to the concentration of NDF and directly related to the rate of particle breakdown of feed and rumen digesta outflow (Kuoppala et al., 2009). Rate of fermentation in the rumen and subsequent particle breakdown are considered to contribute to the rate of rumen clearance (Moseley & Jones, 1984), which in turn influences DMI in legumes (Dewhurst, 2013). Legume leaves are more easily broken down into smaller particles than grass leaves (Mtengeti et al., 1996), and, as a consequence, rumen fermentation is generally increased when red clover silage is offered (Vanhatalo et al., 2009). However, this increase is not always associated with an increase in production of VFAs (Dewhurst et al., 2003b).
In general, evidence within the literature suggests that red clover inclusion in the diet increases milk yields (Moorby et al., 2009; Dewhurst, 2013), with this normally attributed to higher DMIs. In agreement with these findings, over the entire experimental period, milk yield tended to be higher with RCGS. However, this overall effect masks a number of underlying trends and effects within individual harvests. For example, there was a trend for the milk yield advantage to increase with increasing red clover content in the silages (P = 0.228, 0.093 and 0.002 for H1, H2 and H3, respectively), with a difference of 2.9 kg/d in favour of RCGS at H3. The absence of a milk yield benefit with RCGS at H1, despite the significantly higher intakes, may have been due to the low overall CP content in the diet (143 g/kg DM with RCGS, compared to 169 g/kg DM with GS), a reflection of the low application of N fertiliser and the low red clover content. The additional energy deposited as body tissue with this treatment supports the RCGS diet having a low protein content. Despite similar intakes at H3, RCGS produced a significantly higher milk yield than GS. However, fat + protein yield was lower for RCGS than for GS.
Offering RCGS had no effect on milk fat content at H1 and H2, while reducing milk fat content at H3. The poor milk fat response to red clover inclusion is in general agreement with the findings of a number of other studies (Dewhurst et al., 2003a; Moorby et al., 2009; Vanhatalo et al., 2009; Johansen et al., 2017). Milk fat production is associated with the availability of acetic and butyric acid in the rumen, which requires degraded fibre to undergo bio-hydrogenation (Murphy et al., 2000). Low ADF and NDF digestibility leads to a reduction in beta-hydroxybutyrate available for de novo fat synthesis within the mammary gland (Heinriches et al., 1997). Furthermore, PPO lowers rumen bio-hydrogenation of polyunsaturated fatty acids (PUFA), with PUFA known to inhibit milk fat synthesis in the mammary gland. Further, lipids may also be incorporated physically in protein complexes resulting from the action of PPO (Halmemies-Beauchet-Filleau et al., 2012). Digestibility of NDF and ADF was lower with RCGS than with GS at H2 and H3. At H2, intake was higher in RCGS than in GS and, so, despite the lower digestibility of NDF and ADF, intake of digestible fibre in the two treatments would have been similar. In contrast, there being no significant difference in intake between the two silages at H3, the lower digestibility of NDF and ADF with RCGS would explain the lower fat content of the milk in that treatment.
In a meta-analysis of 43 studies comparing grass and legume silages offered to dairy cows, in which 29 involved red clover–grass comparisons, milk protein from perennial ryegrass- and red clover-based diets was 31.9 g/kg and 31.5 g/kg, respectively (Johansen et al., 2018). Similarly, in a review that included eight studies comparing red clover and perennial ryegrass silages, with the exception of one study that showed a slight increase in milk protein content due to red clover silage, milk protein was either unaffected or was higher with GS than with red clover silage (Dewhurst et al., 2003a). The results of the current study are in agreement with those from other studies, with milk protein unaffected at H1, tending to be higher with the GS at H2 and being significantly higher at H3. The effects in the latter harvests may have been due to PPO in red clover catalysing the synthesis of quinones, which complex with, and thereby inactivate, enzymes such as proteases, hence reducing protein availability (Lee, 2014).
Nitrogen and energy utilisation
Effects of diet on N and energy utilisation parameters were inconsistent across harvests. For example, a higher proportion of N intake was excreted as faecal N and a lower proportion as urinary N with the RCGS treatment at H1. However, at H2, there was a trend for urinary N/N intake to be higher with RCGS, while manure N/N intake was higher. No differences were observed at H3. These effects largely reflect the relationships between milk protein yield and DMI, as well as the increasing CP content of the RCGS diet moving from H1 to H3 (Cheng et al., 2011). For example, as the red clover content in the silages increased at H2 and H3, proportionately less N was partitioned to milk in RCGS than in GS. Moreover, findings by Moorby et al. (2009) with treatments involving 3:1 and 3:2 ratios of GS: red clover silage versus GS illustrated that although no differences were found in the overall N balance, apparent partitioning of dietary N into milk was significantly lower in diets with 100% red clover.
There was no clear relationship between rumen degradation (or intestinal digestion) of protein and N utilisation efficiency. For example, a higher content of digestible intestinal protein in RCGS than in GS was associated with a higher proportion of N partitioned to faeces at H1. A higher proportion of ruminal undegraded protein was associated with a lower proportion of partitioned to milk in the same treatment at H3. Faeces have less of an environmental impact than urine as they mineralise more slowly (Haynes & Williams, 1993). Urinary N is rapidly converted to NH3 shortly after it is excreted (Varel et al., 1999) and then to volatile nitrous oxides, which are potent greenhouse gases (Kebreab et al., 2004). In contrast, faecal N is converted to NH3 at a much slower rate and is retained in the soil, contributing to accumulation of soil organic matter (Waghorn, 2008).
While Vanhatalo et al. (2009) found a positive effect on whole-body N balance by partially replacing GS with red clover silage, no such effect was observed in the current study at H1 and H3, in agreement with Bertilsson & Murphy (2003), while the opposite effect was observed at H2. However, as only four animals per treatment were used in the nutrient utilisation studies at each harvest, this was not ideal for identifying differences in N retention. Even with activated PPO (found in fresh samples vs. ensiled), Moorby et al. (2009) found that when feeding fresh red clover with a CP content of approximately 20%, N utilisation by dairy cows was not improved relative to the inactivated PPO. They considered that a CP of 20%, similar to that at H3 in RCGS, was too high for PPO to complex with proteases to have an impact, resulting in less N available for milk protein synthesis but increasing the likelihood of the creation of an environmental burden (Castillo et al., 2000).
Differences in energy utilisation between treatments again largely reflect differences in fat + protein yields, relative to nutrient intake. In addition, ammonia – which results from the additional dietary protein degraded in the rumen – is detoxified in the liver, requiring energy (Canfield et al., 1990), and this may have contributed to the lower milk energy/ME intake with the clover-rich RCGS diets at H3.
System effects and practical considerations
Based on the quantities of herbage ensiled from each sward type and the silage DMIs, one hectare of a red clover–grass mixture was able to provide sufficient silage for 811 “cow feeding days”, while the respective value for GS was 989 “cow feeding days”. Although milk yield during the experimental period was higher with RCGS than with GS treatments (2,220 kg vs. 2,103 kg, respectively), milk fat + protein yield was higher with GS treatments (238 kg vs. 231 kg, respectively). However, while the milk yield was driven in part by the substantially higher intakes with RCGS than with GS at H1, largely due to this being the first harvest following establishment, similar effects would not be expected to arise in subsequent years. Furthermore, limits to red clover persistence also need to be taken into account, with re-seeding of red clover swards normally being required every 3–4 yr. However, one of the main benefits of red clover inclusion in swards is the saving on inorganic N fertiliser, which, in this study, was estimated to be 263 kg N/ha over the course of the season. However, as red clover swards still have a requirement for phosphates and potash, the commonly practised application of organic manures may lead to the crop being oversupplied with N.
Furthermore, the study has clearly demonstrated the very different cow performance responses to red clover inclusion, which can arise between individual harvests within a season, especially during the first full season following establishment, when the relative proportions of the two species changed considerably over the season. Indeed, this issue is rarely identified in published studies as most have involved offering silage from a single harvest. The variability in silage composition between harvests also creates very practical challenges, both commercially and experimentally. For example, the very different forage CP levels with RCGS (increasing from 98 to 216 g/kg DM, between H1 and H3, compared to the increase from 132 to 158 g/kg DM for GS) create real practical difficulties in balancing the protein content of the diets offered. The use of a single concentrate type with both forages and across harvests, as was adopted in the current study, resulted in total ration protein levels being inadequate at H1 and excessive at H3. Similarly, while both swards were harvested on the same date at each of H1–H3 in the current study, given the different growth patterns of the swards, different harvesting dates might have been adopted if a “systems”-type study had been undertaken. However, from an experimental point of view, this would have resulted in swards being harvested and ensiled under different sets of weather conditions, confounding the responses of the sward types. Similarly, while both forage types were wilted for the same duration at each harvest, again differing periods of wilting might have been adopted if weather conditions had allowed. However, periods of rainfall during all three harvests in the current study dictated that longer periods of field wilting were not practical. This demonstrates the difficulty in achieving target DM concentrations with clover-rich swards under typical Irish weather conditions, with this being particularly evident at H3.
Conclusions
This study provides an insight into how the feeding and milk-producing characteristics of silage produced from a sward sown as a red clover–grass mixture changes as the season progresses. While intakes were higher and the milk yields tended to be higher with the RCGS over the entire study period, responses varied considerably between harvests, with this largely reflecting the differences in red clover proportion in the silage and the associated effects on total diet protein content and digestibility. In particular, this study demonstrated that account needs to be taken of the low N content of grass in the red clover–grass mixture, with these differences often ignored in studies when silage from highly fertilised grass is mixed with red clover silage to simulate silage from a mixed sward. Effects on N use efficiency largely reflected the differences in N intakes, relative to milk N output, with this driven largely by differences in the composition of the forages offered.