
INTRODUCTION TO KLEBSIELLA IN THE HOSPITAL AND OTHER ENVIRONMENTAL NICHES
Klebsiella pneumoniae, the type species of Klebsiella, was first described in 1882 by Carl Friedländer as an encapsulated bacillus responsible for pulmonary infections. The bacterium, K. pneumoniae, is a Gram-negative, non-motile member of the Enterobacteriaceae family and is epidemiologically linked to nosocomial and community-acquired (CA) infections. Nosocomial or hospital-acquired (HA) K. pneumoniae is responsible for 3%-8% of all recorded bacterial infections [1] with a higher mortality rate than CA-K. pneumoniae (CA-Kp). HA-K. pneumoniae (HA-Kp) harbours more antimicrobial resistance (AMR) genotypes when compared to CA-Kp. In contrast, CA-Kp isolates have a higher prevalence of virulence factor genotypes, such as rmpA/rmpA2, which encodes capsule regulator genes [2].
In contrast to its recognised threat to human health as a nosocomial risk, Klebsiella species are ubiquitous in nature and found in ecologic niches, such as plants and animals. Although K. pneumoniae has been extensively studied in clinical settings, its role as a hitherto unrecognised zoonosis has been overlooked. An important agent of disease in animals, especially when expressing the hypermucoid phenotype, as observed in California sea lions [3] and African green monkeys [4], K. pneumoniae resides in the gastrointestinal tract of domestic/wild animals and humans alike, and colonises foods, such as various meats. This pathogen is also an important aetiologic agent in clinical cases of mastitis [5]. Environmental niches include water (drinking and surface), waste effluent (municipal sewage and industrial), and vegetation (plants and soil; Bagley [6]). Some species of bats have been reported to carry extended-spectrum ß-lactamase (ESBL) and carbapenemase-producing K. pneumoniae (Table 1). The ease with which K. pneumoniae acquires and transfers AMR-encoding genes (ARGs) across various niches via horizontal gene transfer (HGT) mediated by mobile genetic elements (MGEs) has led to K. pneumoniae being likened to a “canary in the coal mine” [8,19].
The seven phylogroups within the K. pneumoniae species complex, their relevance in cases of human infections, and their detection in other non-human niches.
| KpSC phylogroup | Klebsiella taxa | Clinical relevance | Zoonotic relevance | Reference(s) |
|---|---|---|---|---|
| Kp1 | K. pneumoniae sensu stricto | Human gut1
Human infections1 Nosocomial infections1 Human isolates Brazil3 High prevalence in community & nosocomial isolates Guadeloupe12 | Grey-headed flying foxes (bat)2
Animals (e.g., crabs, mussels, dogs, and horses) Environment (e.g., mangroves, water, and lettuce) 3 African bush elephant, Brazil5 Water, The Netherlands7 Amazon river dolphins8 High prevalence in domestic animals, pigs, poultry, & rivers/natural ponds. Low prevalence in bovine and the environment12 | Wyres et al. [7]1, McDougall et al. [8]2, Morgado et al. [9]3, Furlan et al. [10]5, Rodrigues et al. [11]7, Rocha et al. [12]8, Dereeper et al. [13]12 |
| Kp2 | K. quasipneumoniae subsp. quasipneumoniae | Human gut1
Human infections1 Nosocomial infections1 Low prevalence in community & nosocomial isolates Guadeloupe12 | Environment (Germany & canal water, The Netherlands) 7
Wild animals (foxes & coatis)10 Dogs and elephants11 High prevalence in food-producing animals (pigs, bovines, & poultry) & in vegetables; lower prevalence in domestic animals and in other environmental sources12 | Wyres et al. [7]1, Rodrigues et al. [11]7, de Sousa et al. [14]10, Brisse & Duijkeren [15]11, Dereeper et al. [13]12 |
| Kp3 | K. variicola subsp. variicola | Human gut1
Human infections1 Nosocomial infections1 Human isolates Brazil3 Low prevalence in community & nosocomial isolates Guadeloupe12 | Plant-associated1
Grey-headed flying foxes2 Animals (e.g., Bos indicus), environment (e.g., plants) 3 Food (bananas, Mexico), plants (maize, USA), environment (fungus garden)7 Lignin degradation9 Dogs, birds, monkeys11 High prevalence in bovine, and the environment (soil, rivers/natural ponds, fruits, vegetables, flowering plants, & water catchment). Low prevalence in domestic animals, pigs, & poultry12 | Wyres et al. [7]1, McDougall et al. [8]2, Morgado et al. [9]3, Rodrigues et al. [11]7, dos Santos Melo-Nascimento et al. [16]9, Brisse & Duijkeren [15]11, Dereeper et al. [13]12 |
| Kp4 | K. quasipneumoniae subsp. similipneumoniae | Human gut1
Human infections1 Nosocomial infections1 Human isolates Brazil3 Neonatal outbreak China4 Low prevalence in community & nosocomial isolates Guadeloupe12 | Mosquitos3
Pig farm, China6 Environment (farmland soil, China & Lake Kikker, The Netherlands)7 Wild animals (birds & anteaters)10 High prevalence in dogs, bovine, and soil; low prevalence in cats, pigs, and other environmental sources12 | Wyres et al. [7]1, Morgado et al. [9]3, Perlaza-Jiménez et al. [17]4, Zhao et al. [18]6, Rodrigues et al. [11]7, de Sousa et al. [14]10, Dereeper et al. [13]12 |
| Kp5 | K. variicola subsp. tropica | Human gut1
Low prevalence in Community & Nosocomial isolates Guadeloupe12 | Environment7
High prevalence in Bovine, and the environment (soil, river/natural pond, fruits, vegetables, flowering plants & water catchment). Low prevalence domestic animals, pig, & poultry12 | Wyres et al. [7]1, Rodrigues et al. [11]7, Dereeper et al. [13]12 |
| Kp6 | K. quasivariicola | Human gut1
Human infections1 | Environment (wastewater, UK)12 | Wyres et al. [7]1, Dereeper et al. [13]12 |
| Kp7 | K. africana | Human gut1
Human infections1 | Grey-headed flying foxes, 1st non-human case 20212 | Wyres et al. [7]1, McDougall et al. [8]2 |
[Superscript numbering shown under the clinical and zoonotic relevance columns refers directly to the references included in the final column of Table 1].
Together with studies reporting similarities at the genomic level between clinical and environmental isolates of K. pneumoniae, especially in relation to virulence factors [27–29], it may now be necessary to re-evaluate K. pneumoniae and the broader Klebsiella genus as a hitherto unrecognised zoonosis (Table 2). This bacterium could be considered in the context of One Health by studying the impact of K. pneumoniae on the health of humans, animals, and the environment. The One Health paradigm is a global public health strategy with three axes (human, animal, and environment). The quintessential One Health issue that affects this triad is AMR. Emergence of ARGs can arise following long exposure periods and sub-therapeutic usage of antibiotics in animals, with dissemination of these ARGs to human pathogens or the gut microbiota through food, the environment, and other routes. In addition, human and animal antibiotics are largely similar, allowing for selection and the subsequent transfer of AMR genotypes between humans and animals, either directly or via environmental routes [30].
The nine phylogroups within the K. oxytoca species complex, their relevance in cases of human infections, and their detection in other non-human niches.
| KoSC phylogroup | Klebsiella taxa | Clinical relevance | Zoonotic relevance | Reference(s) |
|---|---|---|---|---|
| Ko1 Ko5 | K. michiganensis | ESßL- producing outbreak in a neonatal unit3
Preterm gut4 Isolated from human blood, Spain and Italy7 | Environment (baby bath drain, detergent bottles, and milk room)3
Isolated from the environment (food, soil, surface water, and sewage sludge) and Giant Panda faeces5 | Chapman et al. [20]3, Chen et al. [21]4, Cosic et al. [22]5, Merla et al. [23]7 |
| Ko2 | K. oxytoca | Human gut – pathobiont causing AAHC1
Human infections – pneumonia, respiratory tract, urinary tract, and skin1 Neonatal nosocomial infections2 Isolated from a human5 | Environment (sinks, wastewater drainage systems, washing machines, and medical solutions) 3
Isolated from the environment (food, soil, and sewage sludge)5 | Herzog et al. [24]1, Singh et al. [25]2, Chapman et al. [20]3, Cosic et al. [22]5 |
| Ko3 Ko9 | K. spallanzanii | Isolated from human peritoneal fluid, France and human urine, Italy7 | Isolated from cow faeces, Italy7 | Merla et al. [23]7 |
| Ko4 | K. pasteurii | Isolated from a human wound and peritoneal fluid, France and human faeces, Italy7 | Isolated from sewage sludge and mouse faeces5
Isolated from soil, cattle faeces, milk, and turtle faeces7 | Cosic et al. [22]5, Merla et al. [23]7 |
| Ko6 Ko7 | K. grimontii | Preterm gut4
Isolated from a human wound, France7 | Isolated from the environment (food, soil, and sewage sludge) and an insect gut5 | Chen et al. [21]4, Cosic et al. [22]5, Merla et al. [23]7 |
| Ko8 | K. huaxiensis | Urinary tract infection, China6
Isolated from human faeces, Italy7 | Isolated from cattle faeces, Italy7 | Hu et al. [26]6, Merla et al. [23]7 |
[Superscript numbering shown under the clinical and zoonotic relevance columns refers directly to the references included in the final column of Table 2].
CURRENT TAXONOMY OF THE GENUS, KLEBSIELLA
Understanding the taxonomy of the genus, Klebsiella, is important to enable accurate identification and description of the epidemiology of these opportunistic pathogens. Whole genome sequencing (WGS) and more advanced biochemical techniques have facilitated clarification of the taxonomy of Klebsiella. Two major subdivisions of Klebsiella are now recognised and denoted Klebsiella pneumoniae species complex [KpSC] and Klebsiella oxytoca species complex [KoSC] [7,31] (Fig 1). WGS-based analysis divided K. pneumoniae into 7 phylogroups, consisting of 5 species with a 95%-96% shared average nucleotide identity (ANI). One of these phylogroups, Klebsiella pneumoniae sensu stricto, was designated Kp1; the remaining phylogroups of KpSC include K. quasipneumoniae subsp. quasipneumoniae (Kp2), K. variicola subsp. variicola (Kp3), K. quasipneumoniae subsp. similipneumoniae (Kp4), K. variicola subsp. tropica (Kp5), K. quasivariicola (Kp6), and K. africana (Kp7)). In total, K. pneumoniae sensu stricto comprises approximately 85% of clinical isolates, and all members of this phylogroup are generally referred to as K. pneumoniae [7]. In addition to K. pneumoniae, other members of the KpSC are opportunistic and virulent bacterial pathogens, such as K. variicola and K. quasipneumoniae subsp. similipneumoniae [32,33]. Furthermore, WGS also showed that KpSC has a 90% shared ANI with the 9 other Klebsiella species, including K. huaxiensis, K. oxytoca, K. michiganensis, K. pasteurii, K. grimontii, K. granulomatis, K. aerogenes, K. spallanzanii, and K. indica [7].
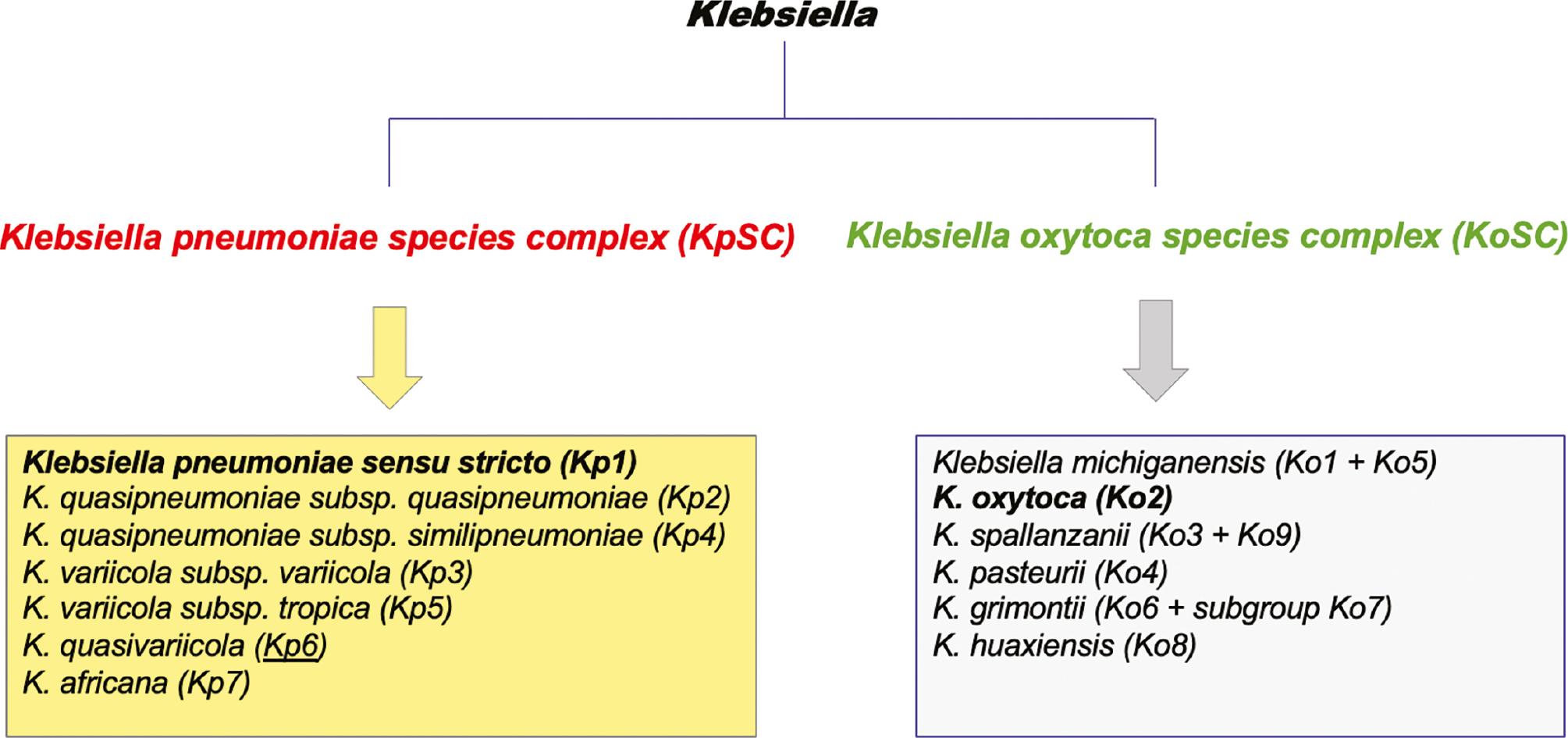
A schematic listing of the taxonomic division of the genus and the members of the K. pneumoniae species complex (KpSC) and the K. oxytoca species complex (KoSC).
The other major subdivision of the genus (Fig 1) relates to the K. oxytoca species complex (KoSC), which consists of the following 6 species: K. michiganensis [Ko1]; K. oxytoca [Ko2]; K. spallanzanii [Ko3]; K. pasteurii [Ko4]; K. grimontii [Ko6]; and K. huaxiensis [Ko8] [31]. These 9 phylogroups are assigned on the basis of variations identified in the sequences of a beta-lactamase-encoding gene (bla OXY). Phylogroups Ko5, Ko7, and Ko9 do not represent a unique species; Ko7 is considered to be a sub-group of Ko6 [22]. Although members of KoSC are less intensively studied, K. oxytoca is the second most likely member of the genus to cause clinical disease after K. pneumoniae [34]. K. oxytoca is an important pathogen and is responsible for antibiotic-associated haemorrhagic colitis (AAHC). Like K. pneumoniae, however, isolates of K. oxytoca have been reported with carbapenem resistance and some shared virulence factors, as discussed below [35]. Representative isolates from the KoSC complex have also been isolated from environmental and animal sources [22,36].
KLEBSIELLA SPECIES CULTURED FROM ANIMALS AND ACROSS THE FOOD CHAIN
The food chain has not traditionally been considered to be a risk factor for transmission of Klebsiella. Despite this, several studies in the recent past have reported isolation of Klebsiella from food-producing animals and food products derived from animals, including milk, salads, and poultry.
Colonisation of domesticated and food-producing/farm animals by Klebsiella species may pose a risk to humans. Co-habiting humans and their companion dogs have been reported to share K. pneumoniae clonal lineages, with some sequence types (STs) identified in dogs having been previously isolated from human infections. Fecal colonisation of dogs by K. pneumoniae was recorded at 39% in one study [37]. A corollary study by the same authors [38] concluded that 60% of K. pneumoniae cultured from companion animals belonged to ST15, which is commonly found in isolates associated with CA- and HA-Kp infections. The majority of these isolates belonged to two PFGE clusters (pulsotypes), which also included human STs.
Contamination of the environment with faeces is another route by which Klebsiella can be transmitted. The faecal-oral cycle was assessed in three dairy farms. The data reported 67% of faecal samples, all of the rumen samples and 89% of water samples obtained were positive for Klebsiella species. Among the genus species detected, K. pneumoniae was the most prevalent in faeces (94%) and rumen (92%), and in swabs obtained from alleyways (100%). Amongst soil and crop samples, K. oxytoca (35% and 7%, respectively), K. variicola (5% and 67%, respectively), and Raoultella planticola (49% and 11%, respectively) were common [39]. Cattle faecal shedding can contaminate the environment and give rise to cases of bovine mastitis, a long-lasting severe intramammary infection that affects milk production. Klebsiella mastitis results in a higher culling rate compared to other forms of mastitis, and this bacterium can express AMR phenotypes. The incidence of Klebsiella mastitis was also reported to be higher between 2010 and 2020 than between 2000 and 2010 [40].
Cases of septicaemia in neonatal pigs have been attributed to specific K. pneumoniae STs, such as ST25, ST278, and ST1978 in Australia and ST25 in England. Pneumonic pigs in Australia were reported to be positive for Klebsiella isolates of ST14 and ST17. Both ST14 and ST1978 have been reported in humans in Australia, and ST25 has been isolated from humans in southeast Asia and Europe [41].
In addition to the threats posed by Klebsiella causing disease in animals, as evidenced by the same STs identified in humans and animals, the bacterium can now be considered to pose a challenge to One Health .
AMR IN KLEBSIELLA SPECIES FROM ANIMALS
The spread of carbapenem-resistant Enterobacteriaceae in the healthcare setting has given rise to concerns about other sectors of the One Health paradigm, such as food-producing and companion animals, and the environment. An investigation of carbapenem-resistant Enterobacteriaceae (CRE) in poultry in China involved the collection of 739 samples from a range of geographic locations and mammals. Some 55 isolates (7.4%) were identified as carbapenem-resistant K. pneumoniae [42]. Further study of these 739 samples in 2019 highlighted 25 K. pneumoniae cultured from chicken (n=19), fly (n=5), and dog samples (n=1), and recorded 22 of the 25 samples as positive for New Delhi metallo-β-lactamase (NDM). These 22 K. pneumoniae samples were comprised of 6 clinically relevant STs, including ST11, ST13, ST37, ST147, ST256, and ST485, and were associated with carriage of bla NDM. Phylogenetic analysis suggested that some of these K. pneumoniae isolates belonging to ST11, ST37, and ST147 were closely related to human isolates. In addition, the most prevalent plasmid replicon types identified were IncFIB [often found in human and animal ESBL-K. pneumoniae], IncX3 [linked to dissemination of bla NDM amongst the Enterobacteriaceae], and IncFII [a carrier of significant ARGs amongst the Enterobacteriaceae] [43].
A recent study in China highlighted 38 publications between 2000 and 2020 that reported on Klebsiella isolated from cases of bovine mastitis and were shown to be resistant to 9 commonly used antimicrobial agents (sulfonamides and tetracyclines). China has banned the use of these two compounds in animal husbandry, suggesting that these Klebsiella species continue to harbour resistance mechanisms [40]. An extensively drug-resistant (XDR) K. quasipneumoniae subsp. similipneumoniae ST5028 was isolated from a Chinese pig farm [18]. A study involving Klebsiella species in Brazil isolated K. quasipneumoniae subsp. similipneumoniae ST1308 in mosquitos, which has also been observed in humans. These Brazilian studies also reported K. pneumoniae ST11, ST15, and ST340 to be present in humans, animals (dogs, crabs, cats, and pigs) and in the environment (water and mangroves). Isolates of sequence types ST437 and ST198 were found in humans and the environment (water and lettuce). K. variicola subsp. variicola contains many STs common across animals, humans, and the environment, with ST355 shared between animals [Bos indicus] and the environment [plants] [9].
The dissemination of ecologically shared STs expressing AMR phenotypes should be monitored in an effort to develop control strategies to mitigate this occurrence.
SPREADING OF MANURE ON AGRICULTURAL SOIL AND CONTAMINATION OF THE ENVIRONMENT BY KLEBSIELLA SPECIES
The spreading of animal manure directly onto farm crops and fields may be a practice that favours the transmission of resistant Klebsiella to plants and the broader environment. Additionally, K. pneumoniae has endophytic abilities, fixing nitrogen in plants, such as alfalfa, maize, and wheat. Because Klebsiella can reside on the stems and leaves of raw produce, K. pneumoniae may become ingested, thus causing infections in susceptible individuals [44–46]. Aquatic environments can also become contaminated following effluent discharges from sources, such as industrial settings, farms, hospitals, and municipal sewage. Bivalve molluscs are used in Norway to assess the level of Klebsiella species in marine environments and to determine the associated AMR phenotypes. Of 476 samples, 204 were confirmed positive for Klebsiella and Raoultella using matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF MS). These samples included K. pneumoniae (n=78), K. oxytoca (n=41), K. variicola (n=33), K. aerogenes (n=1), R. ornithinolytica (n=38), and R. planticola (n=13). An ESBL-producing K. pneumoniae isolate was identified in a marine bivalve growing area that was destined for human consumption, suggesting the food chain as a possible source of transmission to the community [47].
A multicentric study of food products from five European countries reported that one in every two food samples tested was shown to contain a member of the KpSC, and was mainly found in chicken meat and ready-to-eat salads. Kp1 isolates predominated (90.8%) and were found in the tested chicken and salad samples. Kp3 was next highest (6.1%) and shown to be at a much higher rate in salads (18%) than chicken (1%). Similarly, Kp2 (2.3%) was identified in salads alone and Kp4 (0.8%) was found in chicken alone. Identical genotypes found in different food types and in different countries suggested that the potential exists for a high level of transmission across the food chain that could result in human colonisation [48].
It is evident that Klebsiella can be cultured from diverse ecologic niches and that animal and environmental sources can act as a reservoir for the bacterium, where Klebsiella transmission to humans through a combination of direct animal contact or food consumption is currently under-investigated.
ADAPTIVE FEATURES OF KLEBSIELLA ACCOUNT FOR ITS REPERTOIRE OF RESISTANCE GENOTYPES AND VIRULENCE FEATURES
Klebsiella species are known to have an open pangenome that facilitates the acquisition of exogenous DNA, such as various mobile genetic elements (MGEs). This feature enables the bacterium to adapt to the various niches in which it is found. For example, the accessory genome of Klebsiella species aids in the adaptation of the bacterium to ecologic niches by mechanisms that include evasion of human host defence mechanisms. Similarly, AMR and virulence genes are disseminated via MGEs between loci on the genome (insertion sequences, integrons, and transposons) or between bacteria (integrative conjugative elements [ICEs] and plasmids), thus supporting bacterial survival when placed under selection [49]. As a consequence, MDR-hv Klebsiella species resistant to carbapenem-compounds, tigecyclines, and quinolones have emerged. MDR-Kp challenge the ability of clinicians and veterinarians alike to treat these infections.
Important K. pneumoniae virulence factors include the capsule (whose production is regulated by the capsule regulatory rmp-encoding genes) and siderophores (that function to enable iron acquisition: through expression of enterobactin, yersiniabactin, salmochelin and aerobactin; iron transport: ABC-transporter Kfu; and iron suppression: Fur). Lipopolysaccharide [LPS], O-antigens that avoid complement-mediated killing, synthesis is also of importance, as are adhesins, including the type 3 fimbriae for biofilm formation, outer membrane proteins [e.g., ompK genes to protect against neutrophil phagocytosis], type 6 secretion systems [e.g., phospholipase D family protein (PLD1) and one of the type 6 lipase effectors (Tle1), which cause destruction by injecting proteins into target cells] [50]. These features give rise to hv-Kp, a pathotype reported to contribute to increased morbidity and mortality.
In contrast, virulence factors of the KoSC have been less well-studied. Some 14 capsular polysaccharide K-antigens have been identified, with 12 K types being recognised in the KoSC clade and shown to be present in K. pneumoniae (K157 and K164 were identified only in the KoSC). While no LPS O-antigens have been published for the KoSC, seven O types have been identified, all of which have been reported in K. pneumoniae. The only known virulence factor for the KoSC is the AAHC–associated cytotoxin production of tilimycin and tilivalline [51]; however there are some shared virulence factor-encoding genes with K. pneumoniae; including matB, which codes for capsule production, cf29a, fimA, fimH, mrkABCDF, and pilQ, which code for biofilm formation, kfuBC, which codes for iron uptake, and ureA, which codes for urease activity [35].
Biofilm formation is thought to be involved in 65%-80% of bacterial infections reported in the developed world [52,53]. An important virulence feature expressed by K. pneumoniae is its ability to form a thick biofilm on living and abiotic surfaces, such as medical devices, contributing to the XDR phenotype and the severity of infection [54]. K. pneumoniae is a strong biofilm former due its polysaccharide-based capsule, which elaborates a protection mechanism and is important for biofilm surface adhesion and maturation, LPS (initial biofilm attachment and correct folding of type 1 pili), types I and III fimbriae (adhesins that promote binding to living and abiotic surfaces), iron metabolism (siderophore down-regulation to trigger colonisation and infection, as well as immune system evasion), quorum sensing [QS] (types 1 and 2 QS for intra- and inter-species communication for coordinated behaviour of bacterial communities in a biofilm), and its ability to form collaborative biofilms with other bacterial species, such as Pseudomonas aeruginosa and P. protegens [55]. Indeed, vaccines to target these virulence factors have been developed and are in clinical trials [56]. Type III fimbriae have also been detected in K. oxytoca [57], and in AAHC 70% of K. oxytoca isolates are moderate biofilm formers [58].
MDR- and hv-Kp are usually thought of as distinct pathotypes of K. pneumoniae. Based on the comments above, MDR and hv-Kp can be distinguished based on the nature of their genomes. Originally, MDR and hv-Kp were thought to be non-overlapping, but are now being increasingly recognised and converging, thus giving rise to MDR-hv-Kp strains. Bacterial adaptation to any ecologic niche requires the acquisition of genes that confer a selective advantage. Given the open pangenome nature of this bacterium, AMR- and virulence-encoding genes can be acquired and disseminated between Klebsiella to enable this process.
ONE HEALTH AND THE RELEVANCE OF KLEBSIELLA SPECIES
Considering Klebsiella and its ubiquitous distribution across the three domains of One Health , it is not surprising to find that these bacteria are being identified while also being shown to be increasingly resistant to a repertoire of antibiotics (Fig 2). As highlighted by this review, antibiotic selection pressure, interspecies dissemination, and clonal expansion among the Klebsiella strains have been observed, which contributes to the successful spread of Klebsiella-AMR. Antimicrobial agents are used to treat disease in China, but these compounds can also be used to promote growth in large-scale poultry and pig farm production units, with lesser volumes being used in cow and sheep farms. The selective pressure that this imposes reflects the rate of AMR, as reported in one study of 189 K. pneumoniae collected from these 4 species of food-producing animals, in addition to human hospital samples. The rate of MDR-expressing bacteria was shown to be highest (93.6%) among pig isolates, followed by humans (90.4%), chickens (88.9%), cows (52%), and sheep (50%). The susceptibility of these bacteria cultured from these 5 mammalian hosts were tested against a panel of 15 antimicrobial compounds. The data obtained showed that meropenem had the lowest level of resistance recorded (11.6% of isolates were resistant) and ciprofloxacin had the highest level of resistance (77.8% of isolates were resistant). The nosocomial isolates elaborated higher resistance levels to all 15 antimicrobial compounds. Similarly, all 5 animal hosts showed resistance rates > 50% to ciprofloxacin and tetracycline. The prevalence of ciprofloxacin-resistant isolates was higher in chickens (82.2%) and pigs (87.2%), considering fluoroquinolones are widely used in food-producing animals. In contrast, among animal-associated K. pneumoniae, most were susceptible to gatifloxacin, imipenem, and meropenem compounds that are not indicated for use in food-producing animals. Further evidence for the transmission of K. pneumoniae between humans and animals involved the occurrence of ST in animals being detected in hospital settings in China; ST11 was identified in all 5 of the hosts. In addition, ST235 and ST258 were in high abundance in humans, pigs, and chickens [59].
A conceptual schematic describing One Health interconnections that bacteria can harness to disseminate AMR. This was created in Biorender.com.
Colistin, otherwise known as polymyxin E, is an antibiotic that has been clinically reintroduced as a last-resort agent due to the rise in MDR-expressing bacteria. Mobile colistin resistance (mcr) gene acquisition has been reported to be rapidly disseminated by horizontal gene transfer (HGT) among the Enterobacteriaceae. A broad study from a One Health perspective was conducted in which 2855 clinical Klebsiella genomes were screened for all 10 mcr homologs across 5 years in a Chinese hospital (2013-2018). Twenty (0.7%) of these genomes were positive for mcr, of which there were 6 variants (mcr-1.1, mcr-8.1, mcr-8.2, mcr-9.1, mcr-9.2, and mcr-10.1). Both mcr-9- and mcr-10- were susceptible to colistin and mcr-10.1 was discovered in K. pneumoniae, K. quasipneumoniae subsp. quasipneumoniae, and K. variicola. The remaining variants were all found solely in K. pneumoniae. Plasmid typing showed that IncFIIk, IncHI2, IncI2, and IncX4 (in order) were the most common incompatibility groups identified among these variants. Analysis of the mcr-carrying plasmids in GenBank noted highly comparable (> 75% coverage and > 98.5% nucleotide identity) plasmid backbones among the genomes of isolates cultured in chickens, pigs, silver gulls, hospital sewage, and wastewater treatment plants. Because colistin has been more readily used in animal industry than in human healthcare settings, identification of similar mcr variants in humans, animals, and the environment suggests that colistin resistance transmission can be observed in a One Health context. These plasmids also share additional genetic markers, such as other AMR-encoding genes conferring resistance to several classes of antibiotics including those not clinically-associated, a finding that suggests that the occurrence of mcr in these Klebsiella isolates may be due to waste-sources and food-producing animals and not due to clinical colistin selective pressure [60]. Evidence of inter-species and -patient transmission events was observed in a hospital in The Netherlands. In this study 21 isolates from 14 patients were shown to be positive for mcr-1-containing plasmids, including IncX4, IncI2(delta), IncHI2, IncHI2/IncN, and IncHI2/IncQ. These plasmids were found in K. pneumoniae, Escherichia coli, and Kluyvera georgiana, and on the chromosome of K. pneumoniae ST147 [an ST of international high-risk] [61]. These Klebsiella-associated plasmid transmission events carrying the broadest AMR gene repertoires amongst humans, animals, and the environment is an evolutionary process in need of urgent remedial action.
There is a compelling case to be made to monitor Klebsiella in all axes of One Health, including healthcare settings, animals (companion animals, food-producing animals, and exotics), and in the broader environment (e.g. plants and drinking water). To facilitate the initial steps in this process, some useful assembly-based tools for genomic surveillance of K. pneumoniae are already available, including Pathogenwatch, Kaptive, and Kleborate. Pathogenwatch (https://pathogen.watch/) is an online platform that implements genomic analytics for bacterial genera, including Klebsiella. These analytics, including phylogenetics, multilocus sequence typing (MLST), core genome MLST (cgMLST), AMR, virulence gene calling, replicon typing, and identifying loci encoding O- and K-antigens. These features are integrated with epidemiologic data, and visualisation tools provided to aid in global surveillance. Features of Pathogenwatch include the BIGSdb software (http://bigsdb.pasteur.fr/klebsiella/), which can be used to perform MLST and cgMLST, Kleborate (http://github.com/katholt/Kleborate) which is used to identify AMR and virulence loci, O- and K-loci, and wzi genes (the BIGSdb platform screens the genomes against a wzi database to predict K-type), and Kaptive (https://github.com/katholt/kaptive) which performs a more robust capsule and LPS locus typing algorithm [62–65]. Not only will these technologies help elucidate the epidemiology of Klebsiella, but the technologies may also help identify targets for novel control strategies, such as vaccines.
CONCLUSIONS
Our environment has all too often been a neglected component part of One Health [66]. Consequently, when Klebsiella is considered, it is not surprising to find this opportunistic pathogen in the various constituent domains, a feature that consequently may explain why Klebsiella should be regarded as an under-appreciated zoonotic hazard [67]. The recent taxonomic revisions of the KpSC and KoSC clades, together with the lag in development and implementation of accurate diagnostic protocols for this genus, has contributed to this fact. Nonetheless, the extensive antibiotic resistance and hypervirulence phenotypes that can be expressed among isolates of importance to One Health need to be better understood. This feature warrants a reconsideration of how surveillance for this bacterium could be improved, given the evidence presented in this short review.
