
INTRODUCTION
Aspergillus fumigatus is widely found in the environment, particularly in the air, water, plants, and soil, and is the main pathogen causing aspergillosis; A. fumigatus can be isolated from more than 80% of samples from patients with aspergillosis [1]. Conidia of A. fumigatus, owing to their small size (diameter of 2–3 μm), are inhaled by humans and animals and travel deep into the lower respiratory tract; these conidia may be responsible for a variety of clinical cases. However, invasive aspergillosis, which is found primarily in immunocompromised people, remains the most dangerous because of its high mortality rates, which range from 30% to 100% [2–4].
Alveoli are the principal origin of systemic infections of Aspergillus, although infection may also arise from other anatomical sites, such as the sinuses. Alveolar macrophages represent 90% of the resident leucocytes in the lungs, and they home to areas just beneath the alveolar surfactant film [5]. The alveolar macrophages engulf A. fumigatus conidia and subsequently produce cytokines and chemokines, such as TNF-α, IL-12, IFN-γ, IL-18, IL-6, IL-1β, MIP-1, MIP-1α, MIP-2, G-CSF, and GM-CSF [6]. Consequently, professional phagocytes, such as circulating macrophages, neutrophils, monocytes, and dendritic cells, are massively recruited to infection sites. These professional phagocytes play essential roles in host defense against fungal infection through various mechanisms, such as efficiently binding and ingesting A. fumigatus conidia, secreting proinflammatory cytokines, and producing reactive oxygen species [7,8]. Furthermore, damaged tissues release chemokines during inflammation, which attract white blood cells to injury sites [9,10].
Phospholipase D (PLD) is an important phospholipase that catalyzes the hydrolysis of phosphatidylcholine, the most abundant membrane phospholipid, thus producing phosphatidic acid and choline [11]. In mammals, two PLD isoforms, PLD1 and PLD2, are found on the cell membrane surface and are responsible for the phosphatidylcholine-hydrolyzing activity [12]. Host PLD1 and PLD2 are central signaling proteins, and their deficiency in mice leads to complicated developmental, behavioral, and immunological effects, such as defects in T cell signaling and expansion, Fc-mediated phagocytosis in macrophages, and activation of NADPH oxidase in neutrophils [13]. Previous studies have shown that PLD is activated in early stages of β2 integrin-mediated phagocytosis in neutrophils [14], and the two isoforms of PLD—PLD1 and PLD2—coordinately regulate macrophage phagocytosis [15,16] and cell migration [17].
The activity of PLD enzymes in host cells is associated with antimicrobial activities [18]. PLD enzymes in the host can be targeted to halt viral replication during infection. PLDs in macrophages participate in the phagocytosis of Yersinia pseudo tuberculosis and the elimination of Salmonella typhimurium. PLD inhibitors such as VU0285655 block the proliferation of Plasmodium falciparum (a pathogenic factor in malaria) and that of Toxoplasma gondii (the pathogenic factor in toxoplasmosis), on the basis of in vitro experiments. According to our previous research, PLD in lung epithelial cells is activated not only by β-1,3-glucan on the surfaces of A. fumigatus conidia but also by gliotoxin, the most potent toxin produced by A. fumigatus. The activation of both PLD1 and PLD2, two subtypes of mammalian PLD, is important for the efficient internalization of A. fumigatus into lung epithelial cells in vitro [19,20]. However, little is known regarding the consequences of PLD deficiency in the host (through deletion of both the pld1 and pld2 genes) on antifungal ability during A. fumigatus infection in animal models.
In this study, we used pld1-/-pld2-/- mice to analyze the function of PLD during A. fumigatus infection under immunocompetent or immunosuppressed conditions. Deficiency in both pld1 and pld2 genes in mice had only moderate effects on the number of innate immune cells in BALF. Under immunosuppression, PLD deficiency inhibited the production of the cytokine IL-12 and increased production of the anti-inflammatory cytokine IL-10. This response was associated with an increase in A. fumigatus invasion and growth in lung tissue, and faster death in pld1-/-pld2-/- mice than wild type mice.
MATERIALS AND METHODS
A. fumigatus strains and culture
The A. fumigatus wild type strain B5233 was a kind gift from Dr. K. J. Kwon-Chung (National Institutes of Health, Bethesda, Maryland). Conidia were inoculated on Sabouraud dextrose agar and propagated for 5–8 days at 37°C. Then the newly growing conidia were collected with 0.1% Tween-20 and passed through a 40 μm filter to remove hyphal fragments. Finally, the conidia were washed three times in PBS and counted with a hemocytometer before being stored at 4°C.
Mouse infections
All animal studies were approved by the Laboratory Animal Welfare and Ethics Committee (IACUC-13-2016-002) and were conducted according to relevant guidelines and regulations for laboratory animals. C57BL/6 mice were obtained from Fengtai Animal Center of Military Medical Science Academy. pld1-/-pld2-/- mice were obtained from Prof. Dr. Bernhard Nieswandt (University Hospital Wurzburg, Germany). During the normal feeding process, the hair, activity, mental state, balance, and basic vitality pld1-/-pld2-/- mice were comparable to those of wild type mice, and the weights of mice at 10 weeks of age did not significantly differ. With increased age, pld1-/-pld2-/- mice generally became heavier than wild type mice. The invasive pulmonary aspergillosis mouse model was created through modified immunosuppression with steroids, according to a protocol described by Sugui et al. [21]. The dosage and administration of hydrocortisone acetate were as described by Zhang et al. [22]. Image-Pro Express 6.0 was used to process the images (Media Cybernetics Inc., MA, USA). The lungs were homogenized, the homogenates were cultured on Sabouraud dextrose agar medium, and the numbers of colonies were counted after 18 h of culture.
Collection of BALF and alveolar cells
BALF was collected after mice were anesthetized with 25% (v/v) ethyl carbamate intraperitoneally. The lungs were washed eight times with 0.5 ml of PBS, as previously described [6]. The supernatant was frozen at −80°C for subsequent use for the quantification of inflammatory cytokines. All BALF was collected in 50 ml centrifuge tubes and centrifuged for 8 min at 500 g, 4°C. Red blood cells in cell pellets were lysed with erythrocyte lysis buffer. For subsequent flow cytometry, total cell counts in BALF were determined with a Coulter Counter (Cellometer Auto 2000).
Flow cytometry assays
Total cells were collected from BALF in mice and incubated with Fc block anti-mouse CD16/CD32 (Cat:101319), followed by anti-mouse antibodies, including anti-Ly6G-PE (Cat:127608), anti-CD11b-FITC (Cat:101205), and anti-F4/80-PE (Cat:123110), all from BioLegend. Total cells were examined with a BD FACSCanto II flow cytometer and analyzed by FlowJo V10 software.
Multiplex immunoassay for inflammatory cytokines
As previously described, the BALF supernatant was collected from mice at the indicated time points. Multiplex immunoassays were used to determine the concentrations of inflammatory cytokines in the BALF, according to the manufacturer’s instructions (eBioscience).
Statistical analysis
Data are presented as mean ± standard error of the mean for three independent replicates. The significance of differences was assessed with log-rank analysis (for comparative survival), one-way ANOVA, or unpaired Student’s t-tests with a 95% confidence interval in GraphPad Prism software. *p < 0.05, **p < 0.01, and ***p < 0.001.
RESULTS
Deletion of pld1 and pld2 genes increases death and pulmonary fungal burden after A. fumigatus infection in mice with hydrocortisone acetate induced immunosuppression. A. fumigatus is an airborne pathogen that causes life-threatening conditions in patients receiving immunosuppressive therapy or people with congenital neutrophil dysfunction [23]. Immunocompetent mice and pld1-/-pld2-/- mice have been reported not to die after A. fumigatus infection [24]. The immunosuppressed mouse model for the development of invasive pulmonary aspergillosis after A. fumigatus infection was established in the current study with hydrocortisone acetate [21]. The survival rate of pld1-/-pld2-/- mice with A. fumigatus infection was lower than that of wild type mice after immunosuppression with hydrocortisone acetate (Fig 1A); however, loss of pld1 and pld2 genes did not affect the survival rate of immunocompetent mice. In terms of immunosuppression, pld1-/-pld2-/- mice had significantly higher pulmonary fungal burden (Fig 1B), faster growth and invasion of A. fumigatus conidia in lung tissue, and more severe lung tissue damage than wild type mice. Microscopic examination of the lungs of both genotypes of mice after A. fumigatus infection revealed large areas of bronchopneumonia consisting of hyphae surrounded by extensive inflammatory infiltrates, and the pulmonary architecture was effaced by zones of necrosis and inflammation with hemorrhage and edema (Fig 1C). Greater numbers of prominent focal areas of dense inflammatory infiltrates were observed in the lungs of wild type mice than pld1-/-pld2-/- mice; moreover, the necrosis, hemorrhage, and edema in the lung tissue was more severe in pld1-/-pld2-/- mice than wild type mice, and the hyphae in the lung tissue were longer in pld1-/-pld2-/- mice than wild type mice at the same time point after infection (Fig 1C). These data suggested that loss of both pld1 and pld2 genes made immunosuppressed mice more susceptible to A. fumigatus infection, but had less of an effect on the survival rate of immunocompetent mice.
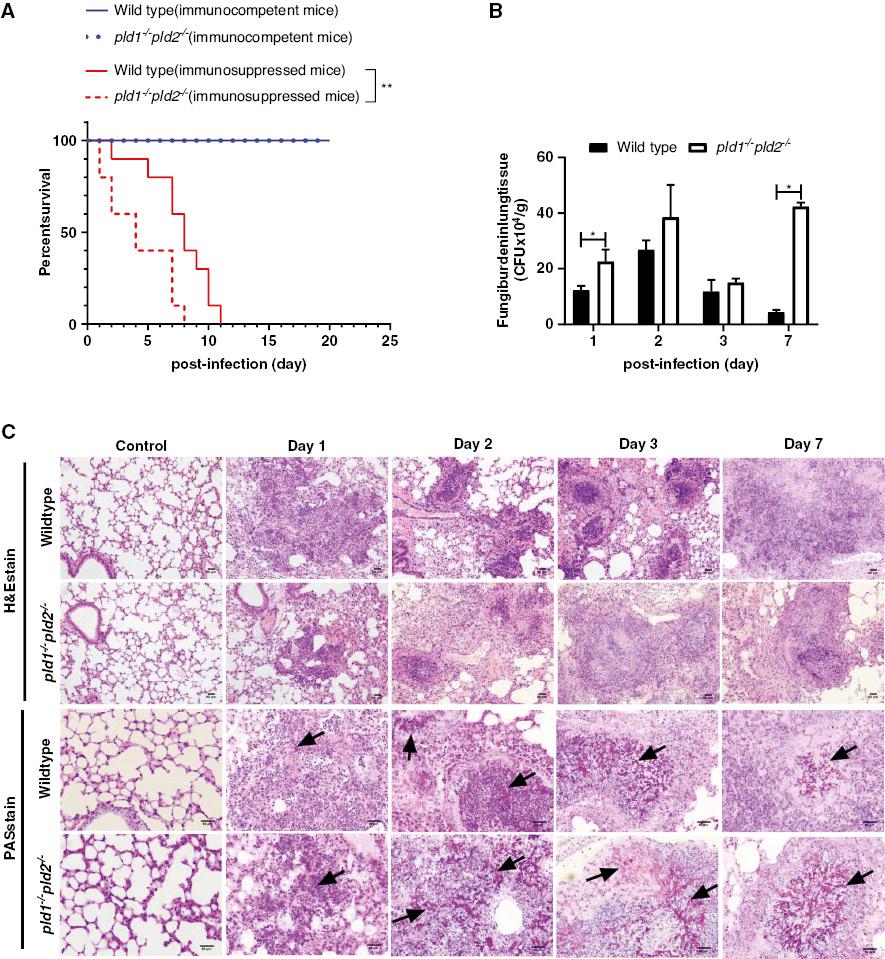
After hydrocortisone acetate induced-immunosuppression, pld1-/-pld2-/- mice die earlier and have higher pulmonary fungal burden than wild type mice after A. fumigatus infection.
Intranasally, the mice were inoculated with 5×106 CFU conidia of A. fumigatus wild type B5233 after being immunosuppressed with hydrocortisone acetate. The immunocompetent mice were inoculated intranasally with 1×107 CFU conidia. (A) For survival analysis, ten mice were weighed every 24 h from the day of infection and were visually inspected twice daily. (B) Mice were sacrificed at post-infection day 0 (control, three mice), day 1 (five mice), day 2 (four mice), day 3 (three mice), and day 7 (three mice). Fungal burden in the lung tissue was measured by counting CFUs. (C) Lung tissues were stained with hematoxylin and eosin (H&E) and periodic acid-Schiff (PAS), and observed by light microscopy (Olympus BX51) at a magnification of 200× or 400×. Scale bar = 50 μm. Black arrows indicate hyphae (red). The images were later processed in Image-Pro Express 6.0 (Media Cybernetics Inc., MA, USA). Three independent experiments with three individual replicates were performed. The significance of differences was assessed with log-rank analysis (for comparative survival) or unpaired Student’s t-tests with a 95% confidence interval in GraphPad Prism 8.0.2 software. * p < 0.05, ** p < 0.01.
Effects of deficiency in both pld1 and pld2 genes on innate immune cell populations in BALF from immunocompetent mice infected by A. fumigatus. Professional phagocytes, such as macrophages, neutrophils, and monocytes, play important roles in preventing fungal infection [5]. Phagocytosis and bactericidal function in these phagocytes are inhibited in vitro when PLD function is deficient [15,25,26]. However, the effect of PLD deficiency on professional phagocytes during A. fumigatus infection was unclear. Here, the cell populations in the BALF from mice were analyzed by flow cytometry with F4/80, CD11b, and Ly6G, which are surface markers of macrophages, monocytes, and neutrophils, respectively.
Alveolar macrophages (F4/80+CD11b-) were the predominant cell populations in BALF from immunocompetent mice before intranasal administration of A. fumigatus conidia. The proportion of alveolar macrophages in pld1-/-pld2-/- mice (93.5%) was not significantly less than that (95.9%) in wild type mice (Fig 2A). At 6 h after administration of conidia, alveolar macrophages remained the major cell populations in BALF from both wild type and pld1-/-pld2-/- mice (Fig 2B). From 0 h to 6 h, the proportions of monocytes (F4/80+CD11b+, approximately seven fold increase, from 0.28% to 1.97%) and neutrophils (Ly6G+CD11b+, approximately three fold increase, from 0.58% to 1.72%) increased significantly in wild type mice, whereas the proportions of neutrophils and monocytes in pld1-/-pld2-/- mice decreased slightly (Fig 2A, 2B). The proportion of neutrophils and monocytes in alveolar lavage fluid from wild type mice was 2.1 times and 1.9 times higher than that from pld1-/-pld2-/- mice, respectively (Fig 2B). At 24 h after conidial administration, alveolar macrophages remained the major cell population in BALF from both wild type and pld1-/-pld2-/- mice (Fig 2C). From 0 h to 24 h, the proportion of alveolar macrophages in wild type mice decreased from 95.9% to 88.2%, and the proportion of alveolar macrophages in pld1-/-pld2-/- mice decreased from 93.5% to 87.7%; however, the proportion of monocytes and neutrophils increased from 0.28% to 4.49% and from 0.58% to 5.18% in wild type mice and pld1-/-pld2-/- mice, respectively (Fig 2A, 2C). Notably, the proportions of the three types of cells were higher in wild type mice than pld1-/-pld2-/- mice (Fig 2C). Moreover, the numbers of neutrophils and monocytes in BALF gradually significantly increased after infection, whereas the number of total cells and macrophages did not change significantly in both genotypes of mice. The numbers of total cells and the three types of immune cells in the pld1-/-pld2-/- mice were slightly lower than those in the wild type mice, but no significant difference was observed between groups (Fig 2D).
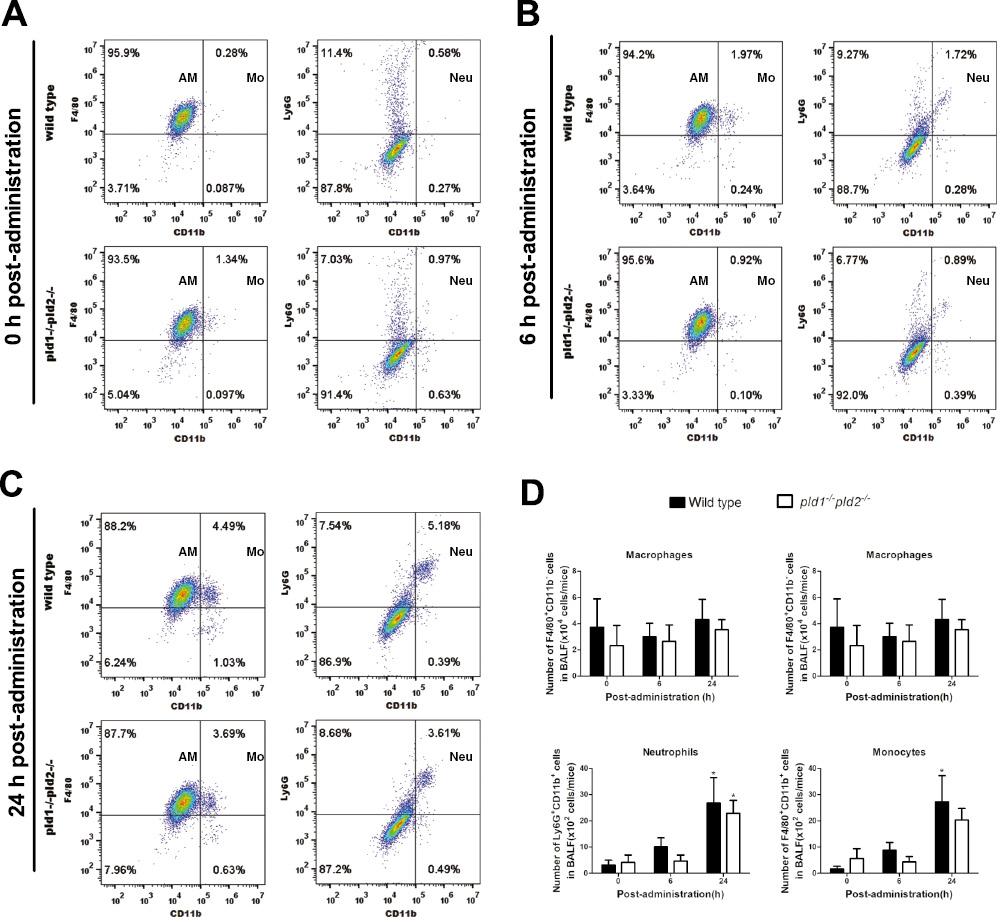
In immunocompetent mice, alveolar macrophages are the major cell populations in BALF after intranasal administration of A. fumigatus conidia.
Immunocompetent mice were inoculated intranasally with 1×107 CFU conidia and anesthetized with 25% (v/v) ethyl carbamate intraperitoneally. BALF was collected at 0 h, 6 h, and 24 h after administration. Each group in each experiment included seven mice. The alveolar lavage fluid of the seven mice was mixed in one 50 ml sterile centrifuge tube. Total cells were collected after centrifugation and incubated with Fc block anti-mouse CD16/CD32 (Cat:101319), followed by anti-mouse antibodies, including anti-Ly6G-PE (Cat:127608), anti-CD11b-FITC (Cat:101205), and anti-F4/80-PE (Cat:123110). (A–C) The content of alveolar macrophages, neutrophils and monocytes in BALF at post-administration 0 h, 6 h and 24 h was examined by BD FACSCanto II flow cytometer and analyzed by FlowJo V10 software. (D) The number of total cells in BALF were determined using a Coulter Counter (Cellometer Auto 2000), and the number of alveolar macrophages, neutrophils and monocytes in BALF was calculated based on total cell number and their respective proportions. Note: The total cells were mixed cells from the alveolar lavage fluid of seven mice in each group. Two independent experiments were performed. p < 0.05, * infected vs. uninfected mice. Abbreviations: AM, alveolar macrophages; Neu, neutrophils; Mo, monocytes.
These findings suggested that macrophages are the major cells in the pulmonary alveoli initially after A. fumigatus infection, and neutrophils and monocytes were recruited into the pulmonary alveoli in response to A. fumigatus infection in immunocompetent mice, but knocking out both pld genes had little effect on the observed cellular recruitment.
Effects of deficiency in both pld1 and pld2 genes on innate immune cell populations in BALF from immunosuppressed mice infected by A. fumigatus. Next, we explored the possible effects of deletion of both pld1 and pld2 genes in immunosuppressed mice on the recruitment of major innate immune cells in the lungs without A. fumigatus infection. The mice were treated with hydrocortisone acetate to establish the immunosuppression model [21]. Alveolar macrophages remained the major cell populations in BALF from both wild type and pld1-/-pld2-/- mice under immunosuppression, but their proportion was lower than that in immunocompetent mice (Fig 3A, compared with Fig 2A). At day 1 post-infection, in BALF from both wild type and pld1-/-pld2-/- mice, the proportion of alveolar macrophages descended sharply, from 76.2% to 0.26% and from 77.9% to 0.58%, respectively; however, neutrophils increased markedly, from 0.29% to 87.0% and from 0.77% to 81.4%, respectively, and became the major cell populations in BALF from both wild type and pld1-/-pld2-/- mice (Fig 3A, 3B and 3D). A consistent decrease in both the proportion (days 1 and 3) and number of neutrophils (day 1) was observed in the pld1-/-pld2-/- mice, as compared with wild type mice (Fig 3B–3D); however, these differences were not significant. In addition, no statistical difference in the number of total cells and the three types of immune cells in BALF was found between groups (Fig 3D).
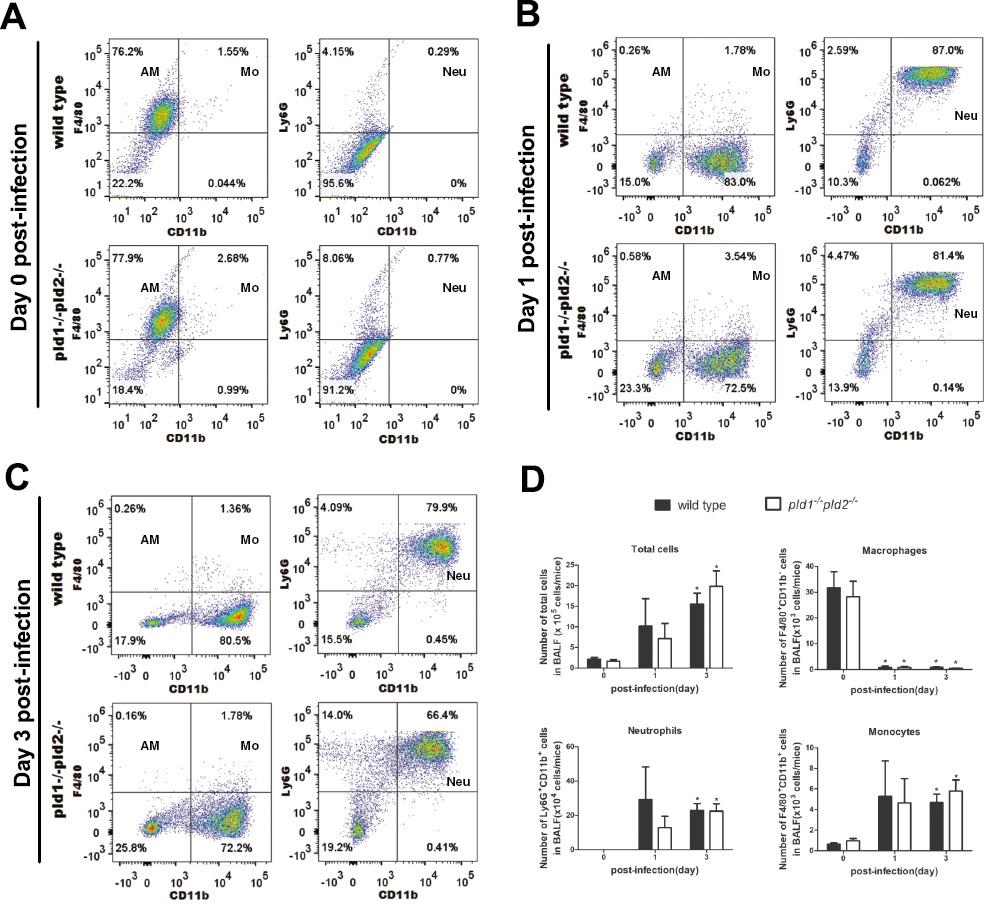
In mice with hydrocortisone acetate-induced immunosuppression, many neutrophils are recruited into BALF and become the major cell populations during A. fumigatus infection.
Mice with immunosuppression were inoculated intranasally with 5×106 CFU conidia and anesthetized by intraperitoneal administration of 25% (v/v) ethyl carbamate. BALF was collected at post-infection day 0, day 1, and day 3. The number of mice in each group ranged from five to seven in each experiment. The alveolar lavage fluid of each group was mixed in one 50 ml sterile centrifuge tube. Total cells were collected after centrifugation and incubated with Fc block anti-mouse CD16/CD32 (Cat:101319), followed by anti-mouse antibodies, including anti-Ly6G-PE (Cat:127608), anti-CD11b-FITC (Cat:101205), and anti-F4/80-PE (Cat:123110). (A–C) The content of alveolar macrophages, neutrophils and monocytes in BALF at post-infection Day 0, Day 1, and Day 3 was examined by BD FACSCanto II flow cytometer and analyzed by FlowJo V10 software. (D) The number of total cells in BALF were determined using a Coulter Counter (Cellometer Auto 2000), and the number of alveolar macrophages, neutrophils and monocytes in BALF was calculated based on total cell number and their respective proportions. Note: The total cells were mixed cells from alveolar lavage fluid of seven, six and five mice in the day 0, day 1, and day 3 post-infection groups, respectively. Three independent experiments were performed. p < 0.05, * infected vs. uninfected mice. Abbreviations: AM, alveolar macrophages; Neu, neutrophils; Mo, monocytes.
These findings indicated that in both genotypes of mice treated with hydrocortisone acetate and infected with A. fumigatus, many neutrophils and some monocytes were recruited into the lungs. However, deficiency in both pld1 and pld2 genes had little effect on the recruitment of neutrophils and monocytes into pulmonary alveoli.
Effects of deficiency in both pld1 and pld2 genes on the release of inflammatory factors in BALF from immunosuppressed mice infected by A. fumigatus. Next, we examined the effects of deficiency in PLD on the release of inflammatory factors in BALF from immunosuppressed mice with A. fumigatus infection. During A. fumigatus infection, the concentrations of the chemokine MCP-1, and the pro-inflammatory cytokines TNF-α, IL-6, and Gro-αKC (IL-8) in BALF increased significantly in wild type and pld1-/-pld2-/- mice; the increase was clearer in pld1-/-pld2-/- mice at day 1 and day 3 after A. fumigatus infection (Fig 4A–D). The concentration of IL-10, an anti-inflammatory cytokine, was higher in pld1-/-pld2-/- mice than wild type mice at day 1 and remained high through day 5 (Fig 4E). Unexpectedly, the concentration of IL-12p40 increased dramatically on day 3 and day 5 in wild type mice but not pld1-/-pld2-/- mice (Fig 4F).
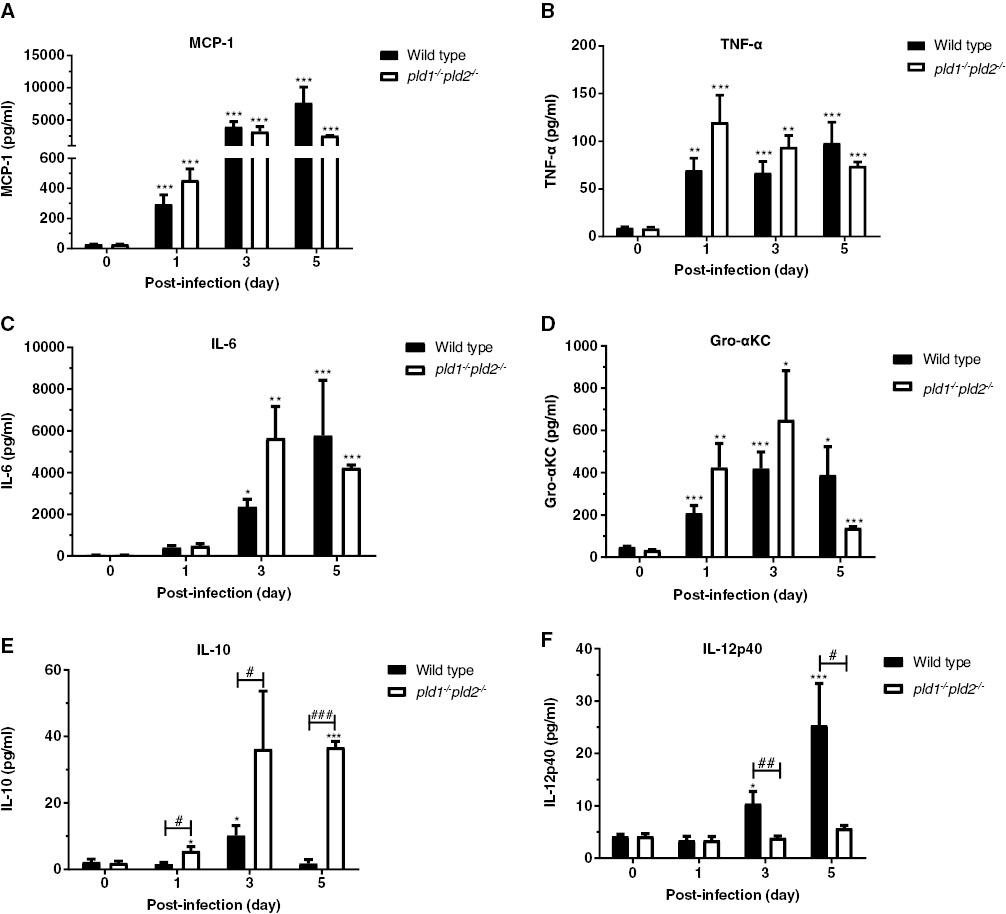
In immunosuppressed mice, deficiency in both pld1 and pld2 genes significantly suppresses the production of IL-12p40 and promotes the production of IL-10 in BALF during A. fumigatus infection.
Mice with immunosuppression were inoculated intranasally with 5×106 CFU conidia and anesthetized by intraperitoneal administration of 25% (v/v) ethyl carbamate. BALF was collected at post-infection day 0, day 1, day 3, and day 5. Each group included six or seven mice. The concentrations of inflammatory cytokines MCP-1 (A), TNF-α (B), IL-6 (C), Gro-αKC (D), IL-10 (E) and IL-12p40 (F) in BALF were detected with multiplex immunoassays according to the manufacturer’s instructions (eBioscience). Statistically significant differences were determined with one-way ANOVA followed by Tukey post-hoc test (vs day 0, ***p < 0.01, **p < 0.01 and *p < 0.05). The significance of differences was determined with unpaired Student’s t-tests with a 95% confidence interval (vs wild type, # p < 0.05, ## p < 0.01, and ### p < 0.001) in GraphPad Prism 8.0.2 software.
These findings suggested that A. fumigatus infection significantly increased the content of the inflammatory factors MCP-1, TNF-α, IL-6, Gro-αKC, and IL-10 in the alveolar lavage fluid of mice under hydrocortisone acetate induced immunosuppression. Deficiency in both pld1 and pld2 genes in mice promoted the release of IL-10 and suppressed the release of IL-12p40 during A. fumigatus infection.
DISCUSSION
Despite being discovered several decades ago, the role of PLD continues to be more precisely elucidated. Most studies have investigated the functions of PLD in cancer, aging-associated diseases, thrombosis, and multiple sclerosis, thus indicating its potential as a target for therapeutic intervention [13]. Most mammalian PLD research has focused on PLD1 and PLD2. Other potential mammalian isoforms have been reported, including PLD3 (also called Sam-9 or HUK4), PLD4, PLD5, and PLD6 (also called mitochondrial PLD, MitoPLD, Zucchini, or Zuc). To date, no studies have been reported whether PLDs in mammalian cells are involved in host antifungal infections. This study presents the first in vivo data on PLD function against Aspergillus infection in mice.
In immunocompetent and immunosuppressive mice, no significant difference was observed in the number of alveolar macrophages in BALF between wild type and pld1-/-pld2-/- mice before and after A. fumigatus infection, thus indicating that the absence of PLD did not alter the number of macrophages in the alveoli. However, in our previous study, the fungal burden in lung tissue and BALF was significantly higher in pld1-/-pld2-/- mice than wild type mice at 6 h post-administration in immunocompetent mice, and the phagocytosis and the fungicidal ability of isolated bone marrow-derived macrophages were significantly lower in pld1-/-pld2-/- mice than wild type mice [24]. In addition, before A. fumigatus infection, the content and number of alveolar macrophages in BALF from immunosuppressed mice were significantly lower than those in mice with normal immune function, but no such changes were observed in neutrophils and monocytes. After A. fumigatus infection, the content and number of alveolar macrophages in BALF sharply decreased, whereas the content and number of neutrophils and monocytes sharply increased in immunosuppressed mice. Moreover, other studies have demonstrated that corticosteroids impair the killing of A. fumigatus conidia by macrophages [18,27,28]. After phagocytosis by macrophages, another important fungal pathogen, Candida albicans, transitions from the yeast to the hyphal stage, and causes macrophage death and fungal escape [29]. Hence, macrophage damage by hydrocortisone acetate and phagocytosis-associated death might possibly explain the rapid and irreversible decrease in alveolar macrophages in BALF in hydrocortisone acetate-immunosuppressed mice during A. fumigatus infection. Moreover, the damage might have contributed to the increased susceptibility observed in the pld1-/-pld2-/- mice in response to A. fumigatus infection.
Neutrophils play critical roles in host defense against invading microorganisms. Previous studies have shown that PLD activation is an early event in β2 integrin-mediated phagocytosis in neutrophils [14], and both PLD1 and PLD2 coordinately regulate cell migration [17]. PLD enzyme activity in neutrophils and leukocytes is associated with antimicrobial functions, such as phagocytosis, chemotaxis, and membrane ruffling [18]. pld1-/- and pld2-/- neutrophils exhibit impaired migration in vitro, but only pld1-/- neutrophils have impaired tissue extravasation [15]. In this study, in both immunocompetent and immunosuppressive states, the number of neutrophils in BALF increased significantly in response to A. fumigatus infection, particularly in mice with immunosuppression induced by hydrocortisone acetate, and the content and number of neutrophils in BALF sharply increased after A. fumigatus infection. However, no significant difference was observed in the number of neutrophils of BALF between wild type mice and pld1-/-pld2-/- mice. Loss of PLD probably did not affect the recruitment of neutrophils to the alveoli in mice during A. fumigatus infection, but instead directly modulated the phagocytosis and fungicidal activity of neutrophils toward A. fumigatus. In mouse models of sepsis, PLD2 in neutrophils is essential for the pathogenesis of experimental sepsis. Bactericidal activity is significantly elevated in pld2-/- mice, and is mediated by increased neutrophil extracellular trap formation and citrullination of histone 3 through peptidylarginine deiminase activation [25]. The exact mechanism of regulation of PLD on the function of neutrophils against A. fumigatus in vivo requires further exploration. Isolating primary neutrophils from pld1-/-pld2-/- mice for further investigation of aspects such as phagocytosis, chemotaxis, and extracellular traps should prove interesting.
In immunocompetent states, after inhalation of A. fumigatus conidia, the content of inflammatory cytokines IL-6, Gro-αKC, and TNF-α in BALF from pld1-/-pld2-/- mice is significantly higher than that in wild type mice, particularly at 6 h after administration [24]. Similarly, we observed that, in immunosuppressive states, the content of these inflammatory cytokines in BALF from pld1-/-pld2-/- mice was also slightly higher than that in wild type mice at day 1 and/or day 3 post-infection, although no statistical difference was observed. This phenomenon might be explained by deficiency in PLD inducing greater secretion of inflammatory cytokines, which in turn recruit immune cells to combat A. fumigatus infection. In contrast, an excessive inflammatory response is also unfavorable to the host and may cause acute inflammatory injury [30,31]. In addition, the cytokine response was likely to be proportional to the higher fungal burden observed in the pld1-/-pld2-/- mice than wild type mice.
IL-12—consisting of two subunits, IL-12p40 and IL-12p35—is an important immunoregulatory cytokine with multiple biological functions. It is produced mainly by phagocytes (monocytes/macrophages and neutrophils) and dendritic cells, in response to pathogens (bacteria, viruses, intracellular parasites, and fungi) [32,33]. IL-12 protects mice against A. fumigatus and C. albicans infection. Moreover, recombinant human interleukin-12 (IL-12) enhances the ability of human monocytes to elicit an oxidative burst and damage the hyphae of A. fumigatus [34]. Another two members of IL-12 family, IL-23 and IL-27, are involved in the response to A. fumigatus infection [32,35]. However, whether loss or abnormal function of host PLD affects the production of IL-12 during A. fumigatus infection is unknown. In this study, the deletion of two major PLDs, PLD1 and PLD2, significantly inhibited the production of IL-12p40 in mice in response to A. fumigatus infection. However, the exact mechanism of this inhibition by PLD on IL-12p40 release during A. fumigatus infection must be further studied.
IL-10, an immunosuppressive and anti-inflammatory cytokine, is essential in maintaining peripheral immunological tolerance and limiting inflammation-associated tissue damage. Blocking IL-10 signaling enhances fungal clearance during cryptococcal infection in the lungs, and IL-10 blockade might be a potential therapy for the treatment of fungal lung infections [36,37]. IL-10 is deleterious during systemic aspergillosis infection, thereby increasing host susceptibility to lethal infection [38]. During A. fumigatus infection, the concentration of IL-10 in the BALF from pld1-/-pld2-/- mice was significantly higher than that from wild type mice; this aspect was likely to be another essential factor in the higher fungal burden and earlier death of pld1-/-pld2-/- mice than wild type mice.
In summary, this was the first study using pld1-/-pld2-/- mice to investigate the function of PLD against fungal infection in either immunocompetent or immunosuppressed mice. The findings indicated that deficiency in both the pld1 and pld2 genes in mice in vivo makes immunosuppressed mice more susceptible to A. fumigatus infection and significantly affects the release of inflammatory factors during A. fumigatus infection.