
Aging is closely associated with a decline in physiologic functions and an increased risk of cancer development. In a recent paper published in Cell Metabolism, Lopez-Otin et al. [1] discussed the highly complex relationship between aging and cancer, further characterizing the aging and cancer with partially overlapping hallmarks by a meta-analysis ( Figure 1 ).
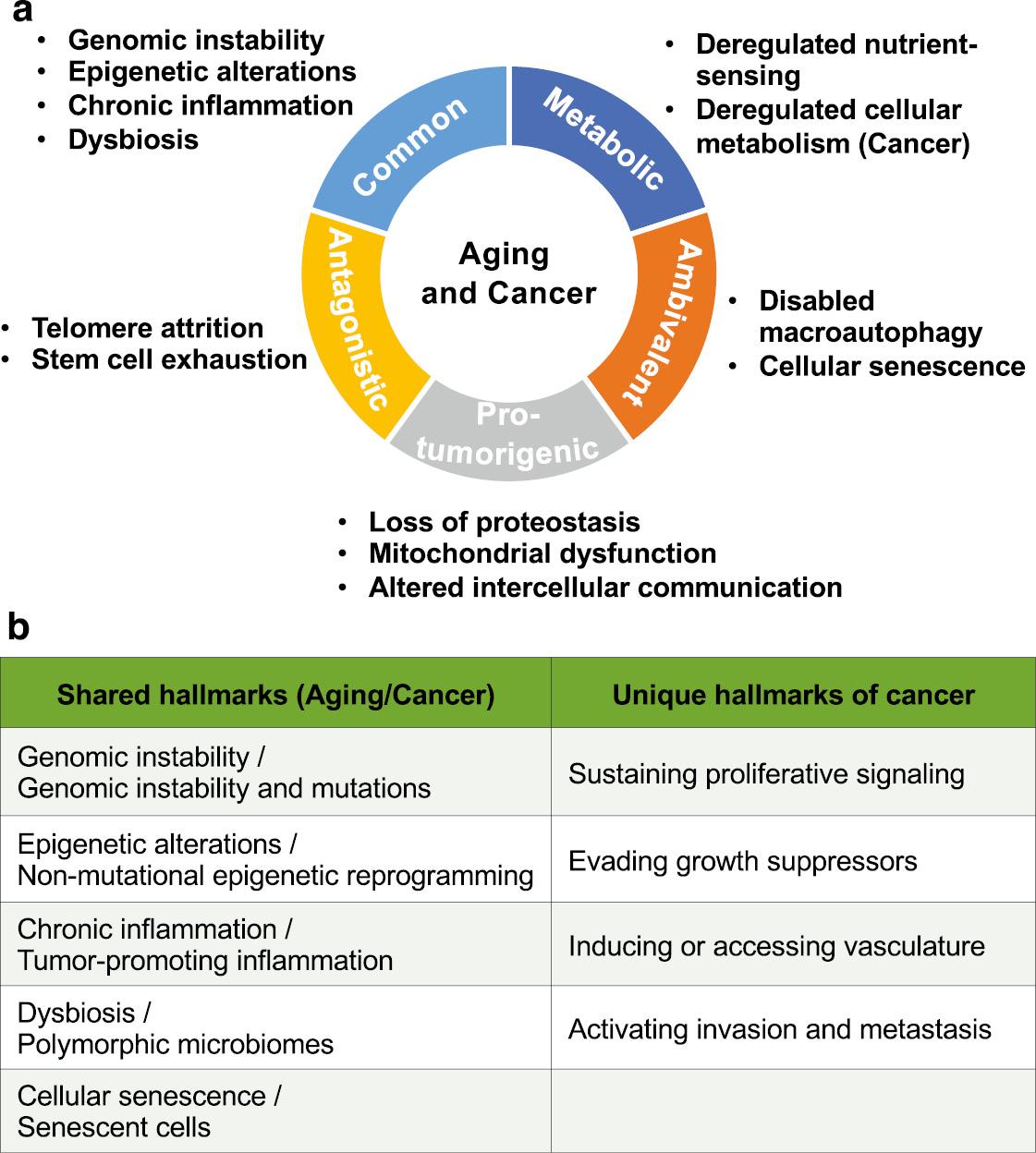
Crosstalk between aging and cancer hallmarks.
(a) Aging hallmarks are categorized into five groups based on correlation with cancer hallmarks. (b) Hallmarks shared between aging and cancer (left column) or as unique to cancer (right column).
Notably, aging is a ubiquitous and multifactorial process that is defined as a time-dependent functional decline in various organisms, including human beings. In 1925, Northrop et al. [2] showed that light intensity affects the growth and lifespan duration in Drosophila, which attracted considerable research interests in aging. Subsequently, aging research gained momentum when longevity mutants were isolated in the nematode, Caenorhabditis elegans, in 1983 [3]. In the past few decades, aging research has become more refined and sophisticated at both the cellular and molecular levels. In 2013 Lopez-Otin et al. [4] summarized the aging process with nine key characteristic features: (1) genomic instability; (2) telomere attrition; (3) epigenetic alterations; (4) loss of proteostasis; (5) deregulated nutrient sensing; (6) mitochondrial dysfunction; (7) cellular senescence; (8) stem cell exhaustion; and (9) altered intercellular communication. In a more recent 2023 review article that examined recent research progress in the aging field, the authors further expanded these fundamental aging hallmarks to involve 12 features, which included 3 additional characteristics: (10) disabled macroautophagy; (11) chronic inflammation; and (12) dysbiosis [5].
An increasing body of evidence has revealed a complex interplay between aging and cancer. To this end, accumulating genetic and epigenetic changes lead to mutations or alterations in gene expression during aging; therefore, aging is a significant risk factor for cancer. In 2000 Hanahan and Weinberg [6] originally classified cancer based on six hallmark features: (1) self-sufficiency in growth signals; (2) insensitivity to anti-growth signals; (3) evading apoptosis; (4) limitless replicative potential; (5) sustained angiogenesis; and (6) tissue invasion and metastasis. In 2011 Hanahan and Weinberg [7] expanded the hallmarks to 10 with 4 additional characteristics: (7) deregulating cellular energetics; (8) avoiding immune destruction; (9) tumor-promoting inflammation; and (10) genome instability and mutation. In 2022 Hanahan [8] further updated the list to 14 characteristics by adding the following: (11) unlocking phenotypic plasticity; (12) non-mutational epigenetic reprogramming; (13) polymorphic microbiomes; and (14) senescent cells. Indeed, the newly updated cancer and aging hallmark features highlight the intensive interplay between these two processes.
Lopez-Otin et al. [1] recently analyzed the partially overlapping hallmark features between aging and cancer ( Figure 1 ), concluding that aging and cancer have a highly complex relationship. More specifically, four aging hallmark features (genomic instability, epigenetic alterations, chronic inflammation, and dysbiosis) are defined almost equally with cancer-promoting factors, including genomic instability and mutations, non-mutational epigenetic reprogramming, inflammation, and polymorphic microbiomes. Therefore, preventive measures targeting these aging hallmarks, such as escaping DNA damage, epigenetic enzyme modulation, anti-inflammatory tablets, and heterochronic fecal microbiota transplantation, will likely be practical measures for cancer prevention. Specifically, the common approaches used to escape DNA damage are quitting cigarette smoking and regularly using UV-blocking sunscreen to prevent lung cancer and melanoma, respectively [9, 10]. Moreover, dietary control may beneficially regulate epigenetic enzyme modulations by supplementation with phytochemicals that have anti-cancer and anti-aging epigenetic effects [11]. The application of fecal microbiota transplantation has also been shown to be a promising approach for preventing colorectal cancer. Fecal microbiota transplantation has also improved the efficacy of cancer immunotherapy for melanoma patients in preclinical studies [12]. In contrast, two aging characteristics, telomere attrition and stem cell exhaustion, likely suppress specific aspects of oncogenesis, and thus can be considered antagonistic hallmark features. Telomerase attrition opposes the replicative immortality of cancer cells, and stem cell exhaustion limits cancer phenotypic plasticity. Therefore, inhibition of telomere maintenance and execution of stem cell exhaustion might be exploited for cancer treatment or prevention. Other aging hallmarks, including loss of proteostasis, mitochondrial dysfunction, and altered intercellular communication, lack an apparent correlation to cancer characteristics, which may promote cell survival, glycolysis, and dysregulated tissue architecture in cancers. Moreover, aging-relevant deregulated nutrient sensing and deregulated cellular energetics in cancer likely exhibit common characteristics involved in metabolic pathways [4]. This commonality partly explains the observations that anabolic signaling promotes aging and cancer, and reduced metabolism prolongs lifespan and inhibits tumor progression.
In addition, disabled macroautophagy and cellular senescence are fundamental yet complex aging features, with both context-dependent tumor-suppressive and tumor-promoting effects. Notably, transient inhibition of autophagy can promote malignant transformation and escape from immunosurveillance; however, autophagy must be reactivated in premalignant cells to improve fitness for subsequent tumor formation [13]. Similarly, acutely-induced cellular senescent cells (acute senescent cells) inhibit cancer formation by a cell-autonomous method or by recruiting T and NK cells. The accumulation of senescent cells (chronic senescent cells) may stimulate inflammation in tissues accompanied by the senescence-associated secretory phenotype (SASP), which consists of proinflammatory cytokines, chemokines, and matrix-remodeling factors [14]. SASP exerts a protective role in tissue regeneration and activation of immunosurveillance to remove senescent cells in acute senescence; however, SASP from chronic senescent cells is harmful and exacerbates senescent status. Therefore, SASP is regarded as a key risk factor for tumorigenesis.
Senescence is largely induced by telomere shortening, which is referred to as replicative senescence, thus leading to irreversible cell cycle arrest in normal cells. Numerous cellular stress stimuli, such as genotoxic, oncogenic, oxidative, and replicative stressors, trigger the onset of premature senescence [15]. Of note, cellular senescence is a hallmark of cancer [8]. Notably, the enhanced oncogenic signaling caused by genetic alterations elicits persistent activation of specific tumor suppressors responsible for monitoring the oncogenic signal intensity. The threshold, which is termed oncogene-induced senescence (OIS), induces cellular senescence [16]. Thus, OIS exerts intrinsic cellular defense mechanisms that prevent cancer development or limit tumor progression. Bypassing OIS by inactivating the tumor suppressor functions is thought to be a major cause of malignant transformation [8]. In addition, cancer cells are subject to the onset of premature senescence triggered by chemotherapy regimens and radiotherapy [17], which is referred to as therapy-induced senescence (TIS). TIS suppresses cell proliferation by inducing a stable cell cycle arrest; however, given the variability of TIS phenotypes, a subset of TIS cells will eventually acquire drug resistance, apoptosis evasion, and dormancy, and ultimately relapse with increased stemness markers. This acquired feature is exploited as a potential therapeutic strategy using senolytic therapy, a selective clearance of senescent cells, which was initially developed to mitigate age-related pathologies and improve longevity [18]. The beneficial senolytic effects have been demonstrated in mice. Specifically, mice that were administered an oral senolytic agent display alleviating aging phenotypes and rejuvenating hematopoietic and muscle stem cells [19]. More importantly, the anti-cancer efficacy of senolytics has been confirmed in phase II studies in combination with multiple senescence-inducing reagents [20]. To this end, sequential drug treatments, in which senescence-inducers are administered first followed by senolytics, exhibit higher effectiveness with reduced toxicity. Senolytic therapy, in contrast, has revealed the heterogeneous nature of senescent cells. As such, different senolytics have variable efficacy among distinct types of senescent cells. In addition, RNA-seq transcriptomic analyses demonstrated the variability of SASP components and transcriptome signatures of core senescent genes among different cell types, stress inducers, and stages post-induction [21]. Considering the heterogeneous nature of senescent cells with accumulating experimental evidence in different cell types or tissues will help identify specific therapeutic target(s) and improve treatment efficacy. Together, cellular senescence, a hallmark of aging and cancer, is a potential therapeutic target of senolytics to treat age-related diseases, as well as therapy-resistant tumors.
Aging is also closely associated with infectious diseases. The recent coronavirus disease 2019 (COVID-19) pandemic has drawn attention to the link between cellular senescence and viral infections [22, 23]. Of note, cellular senescence is induced by viral infections, which is referred to as virus-induced senescence (VIS). VIS is likely caused by infection-evoked cellular stressors, which vary among viruses [23]. Infected and VIS cells release interferons and pro-inflammatory cytokines as part of viral protection mechanisms, thus promoting the paracrine spread of senescence. Specifically, SARS-CoV2-evoked cytokine overproduction and hyperinflammation are thought to facilitate senescence activation in patients infected with COVID-19. In keeping with this notion, elevated senescence markers and increased serum SASP levels have been detected in COVID-19 patients [24]. Notably, administering senolytic agents to SARS-CoV2-infected mice and hamsters eliminated SARS-CoV2-provoked senescent cells. In addition, the agents alleviated COVID-19-reminiscent lung illness and inflammation [24], which suggests that senolytic therapy may be more effective for patients of advanced age. In keeping with this notion, the elderly population shows a higher morbidity and mortality rate for COVID-19, probably because accumulated senescent cells increase SASP secretion, making aged cells more susceptible to VIS and SASP-mediated secondary senescence. These findings suggest that, as with anti-cancer senolytics, anti-infection senolysis is a potentially effective therapeutic strategy for COVID-19 and future pandemics.
In addition to cancer and viral infections, the risk of vascular diseases, which affect the function of blood vessels, also significantly increases with age. Ischemic heart disease ranks as the leading cause of death worldwide, the development and progression of which can be ascribed to cardiovascular cell aging. In 2023 Abdellatif et al. [25] compiled a list of the cardiovascular aging hallmark features by focusing on the mechanisms underlying age-related functional impairments of cardiovascular cells. The hallmark features were categorized into eight sections and the potential of cardiovascular therapeutics that target each hallmark were discussed. Senolytics were also shown to be a promising approach to delaying cardiovascular aging [26].
Taken together, Lopez-Otin et al. classified the convergent, antagonistic, and ambivalent relationships between aging and cancer hallmarks [1]. These intricate connections may lay the theoretical basis for a better understanding of the complex interplay between aging and cancer. More importantly, these mechanistic insights will help determine which strategy is relatively safer in combination with anti-aging or anti-tumor measures for future preclinical research and clinical trials. In addition, cellular senescence, a key hallmark of aging and cancer, has been exploited as a potential senolytic therapeutic target for age-related diseases, therapy resistance cancers, and viral infectious diseases. Hence, further identification of novel aging and cancer hallmarks and analysis of their interconnection will help identify novel therapeutic intervention strategies.